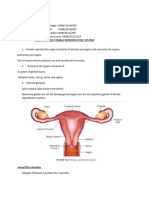
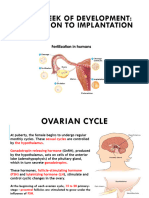
Anda mungkin juga menyukai
- Fisiologi Siklus Haid (DR - Kanadi SumaprajaDokumen70 halamanFisiologi Siklus Haid (DR - Kanadi SumaprajaIntan DhyaksariBelum ada peringkat
- Menstruation & OvulationDokumen42 halamanMenstruation & Ovulationneilesh300Belum ada peringkat
- Fisiologi Siklus HaidDokumen70 halamanFisiologi Siklus HaidsilviBelum ada peringkat
- Cow EstrusDokumen4 halamanCow EstrusMageja TatendaBelum ada peringkat
- Antral Folliculogenesis - ReproductionDokumen19 halamanAntral Folliculogenesis - ReproductionkjhkaBelum ada peringkat
- Normal Menstrual Cycle The ProcessDokumen6 halamanNormal Menstrual Cycle The ProcessBrett StevensonBelum ada peringkat
- Pregnancy Preparation: The Menstrual CycleDokumen4 halamanPregnancy Preparation: The Menstrual CycleTobias GummBelum ada peringkat
- Physiology of OvulationDokumen7 halamanPhysiology of OvulationAza Patullah ZaiBelum ada peringkat
- Menstrual Cycle: Definition of MenstruationDokumen5 halamanMenstrual Cycle: Definition of MenstruationMuhammad Azam Mohd NizaBelum ada peringkat
- UntitledDokumen234 halamanUntitledMarco Antonio AlvarezBelum ada peringkat
- NIH Public Access: Author ManuscriptDokumen13 halamanNIH Public Access: Author ManuscriptDhikaBelum ada peringkat
- Menstrual Cycle Dr. Hani MahdiDokumen80 halamanMenstrual Cycle Dr. Hani MahdiArwa Qishta100% (1)
- Naim LBM 1-1Dokumen38 halamanNaim LBM 1-1dimasBelum ada peringkat
- Obstetrics: The Normal Menstrual CycleDokumen4 halamanObstetrics: The Normal Menstrual Cycleapi-3829364Belum ada peringkat
- Nzary El 3maliDokumen13 halamanNzary El 3maliGana KhaledBelum ada peringkat
- Reproduction (M, F) : Lecturer: Dr. R. Ahangari University of Central Florida, OrlandoDokumen10 halamanReproduction (M, F) : Lecturer: Dr. R. Ahangari University of Central Florida, Orlandokuku411Belum ada peringkat
- The Menstrual Cycle Ob - Gyn SecretsDokumen4 halamanThe Menstrual Cycle Ob - Gyn Secretsleonardotorres1Belum ada peringkat
- OB Dr. Rana (Chapter 5 & 6)Dokumen26 halamanOB Dr. Rana (Chapter 5 & 6)Mich Therese AbejeroBelum ada peringkat
- MK Disorders of Menstruation (OBGY)Dokumen76 halamanMK Disorders of Menstruation (OBGY)Moses Jr KazevuBelum ada peringkat
- FOLLICULOGENESIS4Dokumen14 halamanFOLLICULOGENESIS4Muhammad HamidBelum ada peringkat
- IntroDokumen7 halamanIntroibrahim.21hm32Belum ada peringkat
- Molecular Mechanism of ImplantationDokumen19 halamanMolecular Mechanism of ImplantationAbunu TilayeBelum ada peringkat
- Embryology - Ovarian Cycle: Paper 1 - Part BDokumen9 halamanEmbryology - Ovarian Cycle: Paper 1 - Part BViswa GiriBelum ada peringkat
- MensesDokumen68 halamanMensesdominicamarigu100% (1)
- Stages of Puberty - Sundhed - DKDokumen6 halamanStages of Puberty - Sundhed - DKPavel BerlinschiBelum ada peringkat
- Assignment No 01Dokumen12 halamanAssignment No 01zuha khanBelum ada peringkat
- Female HormoneDokumen43 halamanFemale Hormoneامجد حسين جواد كاظمBelum ada peringkat
- Definition of EmbryologyDokumen12 halamanDefinition of EmbryologyOmar BasimBelum ada peringkat
- 07.12.2019 Human Reproduction-Part 2Dokumen32 halaman07.12.2019 Human Reproduction-Part 2Ayisha AlamBelum ada peringkat
- FOLLICULLOGENESIS DR - SNO, SpOG-KDokumen43 halamanFOLLICULLOGENESIS DR - SNO, SpOG-KEndah PratiwiBelum ada peringkat
- Female Reproductive SystemDokumen3 halamanFemale Reproductive SystemJayricDepalobosBelum ada peringkat
- Fertilization Reflection PaperDokumen2 halamanFertilization Reflection PaperCrisandro Allen Lazo100% (2)
- Menstrual Cycle and Fertility by Neville Mvo NgumDokumen7 halamanMenstrual Cycle and Fertility by Neville Mvo Ngumlovelyc95Belum ada peringkat
- Female Physiology 1Dokumen2 halamanFemale Physiology 1JayricDepalobosBelum ada peringkat
- Physiology of Female Reproductive System The Ovarian CycleDokumen9 halamanPhysiology of Female Reproductive System The Ovarian CycleAmiraah MasriBelum ada peringkat
- JurnalDokumen4 halamanJurnalS1kebidanan Semester 3Belum ada peringkat
- Physiology of The Menstrual Cycle 2Dokumen22 halamanPhysiology of The Menstrual Cycle 2anoy judeBelum ada peringkat
- OvariesDokumen5 halamanOvariesarpitsonone16Belum ada peringkat
- MODULE 1-Lesson 2-3Dokumen11 halamanMODULE 1-Lesson 2-3JOHN BRAINARD PEJOBelum ada peringkat
- Follicular PhaseDokumen9 halamanFollicular PhaseDista ArifanyBelum ada peringkat
- The Menstrual Cycle P. Mock: Infertility and Gynecologic Endocrinology Clinic Geneva University HospitalDokumen12 halamanThe Menstrual Cycle P. Mock: Infertility and Gynecologic Endocrinology Clinic Geneva University HospitalMubashir's iPhoneBelum ada peringkat
- Problem No.1Dokumen34 halamanProblem No.1ayesha bgBelum ada peringkat
- Chapter - 3 Human Reproduction - WatermarkDokumen23 halamanChapter - 3 Human Reproduction - WatermarkDurgesh SainiBelum ada peringkat
- Menstrual Physiology, Amenorrhea: Page 1 of 5Dokumen5 halamanMenstrual Physiology, Amenorrhea: Page 1 of 5seth10Belum ada peringkat
- 4. week ovulation, fertilization, implantation (١)Dokumen41 halaman4. week ovulation, fertilization, implantation (١)Maisy JohnBelum ada peringkat
- ENDO #6 - Reproductive Endocrinology of The Female - 13.10.23Dokumen22 halamanENDO #6 - Reproductive Endocrinology of The Female - 13.10.23sbobine.imsBelum ada peringkat
- Ovo LationDokumen5 halamanOvo LationAsif Hasan NiloyBelum ada peringkat
- Female Reproductive CycleDokumen4 halamanFemale Reproductive CycleTRASH MAILBelum ada peringkat
- CH 5 Implantation, Placental Dev (Part1)Dokumen36 halamanCH 5 Implantation, Placental Dev (Part1)Tengku Chairannisa PutriBelum ada peringkat
- Physiology of Female Reproductive SystemDokumen10 halamanPhysiology of Female Reproductive Systemhamadimariam10Belum ada peringkat
- Physiology of MenstruationDokumen33 halamanPhysiology of MenstruationDebajyoti DasBelum ada peringkat
- Mammalian Reproductive SystemDokumen50 halamanMammalian Reproductive SystemLeena MuniandyBelum ada peringkat
- Embryology 2Dokumen40 halamanEmbryology 2Sri Ram 07Belum ada peringkat
- Imported FileDokumen18 halamanImported Fileg8pn6pt8nhBelum ada peringkat
- Menstrual CycleDokumen41 halamanMenstrual CyclesharimileeratnamBelum ada peringkat
- Physiology of Female Reproductive SystemDokumen16 halamanPhysiology of Female Reproductive Systemsami loveBelum ada peringkat
- Menstrual CycleDokumen22 halamanMenstrual CycleDaniel AtiehBelum ada peringkat
- Hormonal Cycle Lesson PlanDokumen7 halamanHormonal Cycle Lesson Plannisha kaushik100% (2)
- Menstrual Cycle and Its Physiology Mechanism Pathophysiology and Clinical SignificanceDokumen2 halamanMenstrual Cycle and Its Physiology Mechanism Pathophysiology and Clinical SignificanceRoyd KabweBelum ada peringkat
- Introduction to Clinical Reproductive EndocrinologyDari EverandIntroduction to Clinical Reproductive EndocrinologyPenilaian: 1 dari 5 bintang1/5 (1)
- Overview of Cell Cycle by Javali.GDokumen15 halamanOverview of Cell Cycle by Javali.GJavali.GBelum ada peringkat
- Urologic Principles and Practice 2nd EditionDokumen692 halamanUrologic Principles and Practice 2nd EditionRonuve RonuveBelum ada peringkat
- NEET PG 2013 Question Paper With SolutionsDokumen2.164 halamanNEET PG 2013 Question Paper With SolutionsSK RaulBelum ada peringkat
- Biology 209 Lab ManualDokumen128 halamanBiology 209 Lab ManualikomalBelum ada peringkat
- Sports Medicine and Health ScienceDokumen9 halamanSports Medicine and Health ScienceJavier Estelles MuñozBelum ada peringkat
- CSI Physical ExamDokumen3 halamanCSI Physical ExamSympaschoBelum ada peringkat
- 2.2 - Powerpoint D - Effects of Exercise On The Cardiovascular SystemDokumen23 halaman2.2 - Powerpoint D - Effects of Exercise On The Cardiovascular SystemAshraf ShaikhBelum ada peringkat
- Pericardial Disease 1Dokumen100 halamanPericardial Disease 1Anab Mohamed Elhassan Abbas AlnowBelum ada peringkat
- Sleep Phylogeny & OntogenyDokumen7 halamanSleep Phylogeny & OntogenyVivian PBelum ada peringkat
- HURST NCLEX Review Facts PDFDokumen6 halamanHURST NCLEX Review Facts PDFMikeBelum ada peringkat
- CRB Aranceles Ambulatorio 2017bDokumen28 halamanCRB Aranceles Ambulatorio 2017bElizabeth Arancibia NacarateBelum ada peringkat
- Ronald John Recio, M.A., Rpsy, Emdrprac. Mbpss Pamantasan NG Lungsod NG Maynila Puso MoDokumen22 halamanRonald John Recio, M.A., Rpsy, Emdrprac. Mbpss Pamantasan NG Lungsod NG Maynila Puso MoChunelle Maria Victoria EspanolBelum ada peringkat
- Cell Eukaryote Prokaryote Animal Plant 1.: Learning Packet No.2: Prokaryotic and Eukaryotic CellsDokumen3 halamanCell Eukaryote Prokaryote Animal Plant 1.: Learning Packet No.2: Prokaryotic and Eukaryotic CellsSean LestherBelum ada peringkat
- Insecticide ClassificationDokumen10 halamanInsecticide ClassificationAnuragBhatnagarBelum ada peringkat
- Concept Map TemplateDokumen4 halamanConcept Map TemplateaamenaBelum ada peringkat
- Recent Advances in HypertensionDokumen6 halamanRecent Advances in Hypertensionrudi saputraBelum ada peringkat
- ThreePointer's Guide To Histology Steeple Chase For 022 ClassDokumen54 halamanThreePointer's Guide To Histology Steeple Chase For 022 ClassNnaemeka Neboh100% (1)
- HematologyDokumen38 halamanHematologyNicole Ann RoseteBelum ada peringkat
- Dopamine Receptor Subtypes, Physiology and Pharmacology - New Ligands and Concepts in SchizophreniaDokumen17 halamanDopamine Receptor Subtypes, Physiology and Pharmacology - New Ligands and Concepts in SchizophreniaolivukovicBelum ada peringkat
- Skema Jawapan Biologi Percubaan SPM 2008: Structured Question: No. 1Dokumen13 halamanSkema Jawapan Biologi Percubaan SPM 2008: Structured Question: No. 1afifasolehahBelum ada peringkat
- 0612-88-510 Balanced Surg TechDokumen60 halaman0612-88-510 Balanced Surg TechAnonymous a3JsNXNBelum ada peringkat
- Flight PhysiologyDokumen5 halamanFlight PhysiologyRicardo MarmolBelum ada peringkat
- Pelvic Organ ProlapseDokumen68 halamanPelvic Organ ProlapseAlyanna CrisologoBelum ada peringkat
- Pro Fitness Home Gym: Assembly & User Instructions - 237/4693Dokumen36 halamanPro Fitness Home Gym: Assembly & User Instructions - 237/4693Aneta KochańskaBelum ada peringkat
- 2.2 Drug Absorption From The GITDokumen15 halaman2.2 Drug Absorption From The GITCari MosehuusBelum ada peringkat
- Mark Scheme: Double Award Science BiologyDokumen9 halamanMark Scheme: Double Award Science BiologyDaniel LoughreyBelum ada peringkat
- 23312-02 - Dissertation ETH Zürich Michael EderDokumen108 halaman23312-02 - Dissertation ETH Zürich Michael EderMichael EderBelum ada peringkat
- IyotDokumen7 halamanIyotMonaimLaraMacabangonIVBelum ada peringkat
- IB Biology HL SyllabusDokumen9 halamanIB Biology HL SyllabusjessicaBelum ada peringkat
- 2ND Activity Frezel EjemparDokumen5 halaman2ND Activity Frezel EjemparRONALD ARTILLEROBelum ada peringkat