Anda mungkin juga menyukai
- Clase 8 EA Evaluación Nutricional.2010Dokumen25 halamanClase 8 EA Evaluación Nutricional.2010Amelia PomaBelum ada peringkat
- CASO CLÍNICO Cirrosis DescompensadaDokumen16 halamanCASO CLÍNICO Cirrosis DescompensadaDiana MaytaBelum ada peringkat
- MecanismosHambreSaciedadDokumen4 halamanMecanismosHambreSaciedadDayraCampoverde50% (2)
- Receptores GLUT y su función en el transporte de glucosaDokumen1 halamanReceptores GLUT y su función en el transporte de glucosaFernanda UrbanBelum ada peringkat
- Alimentación en El AdolescenteDokumen9 halamanAlimentación en El AdolescenteAna Claudia Bullon Cuadra100% (1)
- Isiologia Del Metabolismo de La Proteinas PDFDokumen19 halamanIsiologia Del Metabolismo de La Proteinas PDFRosario Laguna100% (1)
- Expo ProteinasDokumen15 halamanExpo ProteinasDiegoMauricioGonzalezBelum ada peringkat
- Semiologia Nutricional .PT - EsDokumen26 halamanSemiologia Nutricional .PT - EsGabriela Soto DiazBelum ada peringkat
- Cuestionario Par-Q 2023Dokumen1 halamanCuestionario Par-Q 2023CHRISTOPHER MONTES ALMAOBelum ada peringkat
- Nutricion ParenteralDokumen8 halamanNutricion ParenteralGeancarlos MejiaBelum ada peringkat
- Prescripción NutriciaDokumen2 halamanPrescripción NutriciaErick HernandezBelum ada peringkat
- Caso clínico nutricionalDokumen7 halamanCaso clínico nutricionalAngie TobarBelum ada peringkat
- Dietas vegetarianas: beneficios y riesgosDokumen13 halamanDietas vegetarianas: beneficios y riesgosPierina TorrejonBelum ada peringkat
- Farmacología en Nutrición Unidad IDokumen4 halamanFarmacología en Nutrición Unidad IAlma Guadalupe Marín MagañaBelum ada peringkat
- Farmacos para La AnemiaDokumen10 halamanFarmacos para La Anemiascid100% (1)
- Plan de Atención NutricionalDokumen36 halamanPlan de Atención NutricionalRamón GómezBelum ada peringkat
- FormulasDokumen6 halamanFormulasEdwardwwBelum ada peringkat
- Fisiología Gastrointestinal TripticoDokumen2 halamanFisiología Gastrointestinal TripticoGrace Garcia100% (1)
- Elaboracion de Una DietaDokumen5 halamanElaboracion de Una DietaErnesto LopezBelum ada peringkat
- Dieta MediterráneaDokumen6 halamanDieta MediterráneaCintia VentocillaBelum ada peringkat
- Fases de La DigestionDokumen27 halamanFases de La DigestionDrossxBelum ada peringkat
- MACRONUTRIENTESDokumen36 halamanMACRONUTRIENTESShirley Hinostroza ChamorroBelum ada peringkat
- Act 10 Aciduria Glutárica Tipo 1-Alim. y Nut. en Circunstancias EspecialesDokumen20 halamanAct 10 Aciduria Glutárica Tipo 1-Alim. y Nut. en Circunstancias EspecialesPaola Bejarano100% (1)
- Protocolo Nutricion Parenteral UCIPDokumen15 halamanProtocolo Nutricion Parenteral UCIPHenry LeonardoBelum ada peringkat
- Manual DietologiaDokumen152 halamanManual DietologiaKaren CarreraBelum ada peringkat
- Síndrome de Intestino CortoDokumen2 halamanSíndrome de Intestino CortoJoel Monja Salas100% (2)
- Exploración Física y Medidas AntropométricasDokumen9 halamanExploración Física y Medidas AntropométricasMiguel AcevedoBelum ada peringkat
- Alimentación PediatricaDokumen17 halamanAlimentación PediatricaGabriela Orozco LopezBelum ada peringkat
- Alimentos Ricos en Ácidos Grasos Esenciales - IDokumen6 halamanAlimentos Ricos en Ácidos Grasos Esenciales - IJosé Antonio García-Olveros GonzálezBelum ada peringkat
- Clase 4 Nutrición y Sistema InmunitarioDokumen58 halamanClase 4 Nutrición y Sistema InmunitarioNayadee Javiera Souto CastilloBelum ada peringkat
- Dietoterapia BásicaDokumen36 halamanDietoterapia BásicaIndirita Badillo Alvarado100% (2)
- Nutriogenomica PDFDokumen24 halamanNutriogenomica PDFChristopher Carranza100% (1)
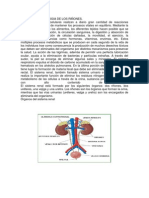
- Anatomia y Fisiologia de Los RiñonesDokumen17 halamanAnatomia y Fisiologia de Los RiñonesArturo Merida RodriguezBelum ada peringkat
- Nutrición para AdolescentesDokumen6 halamanNutrición para AdolescentesCenaidaBelum ada peringkat
- Somatotipo y SomatocartaDokumen3 halamanSomatotipo y SomatocartaAndres BravoBelum ada peringkat
- Indicadores ABCD Necesarios para El Proceso de Atención Nutricia en La Población InfantilDokumen6 halamanIndicadores ABCD Necesarios para El Proceso de Atención Nutricia en La Población InfantilKarla RuizBelum ada peringkat
- Funciones Fisiológicas Del AguaDokumen4 halamanFunciones Fisiológicas Del AguaPaul GarciaBelum ada peringkat
- Nutrición en El Adulto MayorDokumen9 halamanNutrición en El Adulto MayorAmos Instituto Superior en Salud100% (1)
- 19 Interrelaciones MetabólicasDokumen37 halaman19 Interrelaciones MetabólicasLuis VizcayaBelum ada peringkat
- Dieta South BeachDokumen37 halamanDieta South BeachJuan DelgadilloBelum ada peringkat
- Dietoterapia en Pacientes Con Cancer.Dokumen35 halamanDietoterapia en Pacientes Con Cancer.Yanfred Yeferson BernacheaBelum ada peringkat
- Norma manejo malnutrición niños menores 6 añosDokumen36 halamanNorma manejo malnutrición niños menores 6 añosKatty PerezBelum ada peringkat
- FENILCETONURIADokumen30 halamanFENILCETONURIAion sirbuBelum ada peringkat
- El Sistema Digestivo en El NiñoDokumen3 halamanEl Sistema Digestivo en El NiñoAna MalaisiBelum ada peringkat
- Propiedades Generales de Los Hidratos de CarbonoDokumen4 halamanPropiedades Generales de Los Hidratos de CarbonoEdbo 98Belum ada peringkat
- 04, Generalidades de Enf. AutoinmunesDokumen54 halaman04, Generalidades de Enf. AutoinmunesÐiegoFernandoSuarez100% (1)
- Aprendizaje autogestivo en ambientes virtualesDokumen18 halamanAprendizaje autogestivo en ambientes virtualesJose Alberto NHBelum ada peringkat
- Motilidad DigestivaDokumen16 halamanMotilidad DigestivatrumandemontesBelum ada peringkat
- Metabolismo de Los Carbohidratos en El DeportistaDokumen13 halamanMetabolismo de Los Carbohidratos en El DeportistaWilson JavBelum ada peringkat
- Enfermedad de WolmanDokumen3 halamanEnfermedad de WolmanBrayan BennettBelum ada peringkat
- Métodos de Valoraciones DietéticosDokumen1 halamanMétodos de Valoraciones DietéticossaraBelum ada peringkat
- CarbohidratosDokumen8 halamanCarbohidratosHugo Glez PradoBelum ada peringkat
- Fórmulas infantiles: composición, indicaciones y tiposDokumen72 halamanFórmulas infantiles: composición, indicaciones y tiposMaría Manuela BarreiroBelum ada peringkat
- MEGAREXIADokumen8 halamanMEGAREXIAAdrianaJuradoBelum ada peringkat
- Agua, electrolitos y nutriciónDokumen58 halamanAgua, electrolitos y nutriciónSharon Cubas QuezadaBelum ada peringkat
- Caso Clinico GastroenterologiaDokumen23 halamanCaso Clinico GastroenterologiaAlexandra Pauccar Rendon100% (1)
- DIETOTERAPIA 2aDokumen19 halamanDIETOTERAPIA 2aMario MoralesBelum ada peringkat
- 2 5 Indicadores BioquimicosDokumen48 halaman2 5 Indicadores BioquimicosJasmany Sergio Sánchez Aguirre100% (1)
- Nutriciòn e InmunonutriciónDokumen26 halamanNutriciòn e InmunonutriciónMagdalena Rivera BurgosBelum ada peringkat
- Integracion MetabolicaDokumen21 halamanIntegracion Metabolicajuan100% (2)
- Patologias PleuralesDokumen25 halamanPatologias PleuralesAugusto Suárez GómezBelum ada peringkat
- AsmaDokumen15 halamanAsmaAugusto Suárez GómezBelum ada peringkat
- El Papel Evolutivo de La Biopsia de Mama Asistida Por VacíoDokumen10 halamanEl Papel Evolutivo de La Biopsia de Mama Asistida Por VacíoAugusto Suárez GómezBelum ada peringkat
- Asma BronquialDokumen3 halamanAsma BronquialAugusto Suárez GómezBelum ada peringkat
- Dokumen - Tips Informe Tecnico VerificadorDokumen1 halamanDokumen - Tips Informe Tecnico VerificadorElisa PinkBelum ada peringkat
- Corticosteroides TopicosDokumen3 halamanCorticosteroides TopicosAugusto Suárez GómezBelum ada peringkat
- Herencia Mendeliana, Diapos !!Dokumen26 halamanHerencia Mendeliana, Diapos !!Augusto Suárez GómezBelum ada peringkat
- Metabolismo específico de tejidos: hígado, grasa, músculoDokumen62 halamanMetabolismo específico de tejidos: hígado, grasa, músculoJunior Uriarte DávilaBelum ada peringkat
- Ciclo de Cori: la circulación de glucosa y lactato entre músculo y hígadoDokumen7 halamanCiclo de Cori: la circulación de glucosa y lactato entre músculo y hígadoJeanette PaulinaBelum ada peringkat
- Sistemas energéticos en el ejercicio físicoDokumen32 halamanSistemas energéticos en el ejercicio físicoronnie david lopez perezBelum ada peringkat
- Ayuno, glucosa, cetonasDokumen1 halamanAyuno, glucosa, cetonasCristhian Penagos ParraBelum ada peringkat
- Cómo Funciona El Ciclo de CoriDokumen2 halamanCómo Funciona El Ciclo de CoriMatías Córdoba100% (1)
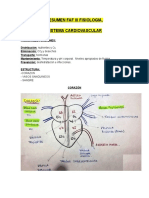
- Resusmen Faf III FisiologiaDokumen17 halamanResusmen Faf III FisiologiamartinBelum ada peringkat
- FISIOLOGIADokumen5 halamanFISIOLOGIAAngelica GalvisBelum ada peringkat
- Tema 6 - GluconeogenesisDokumen23 halamanTema 6 - GluconeogenesisAlejandro Lopez VillanuevaBelum ada peringkat
- Determinación glucosa eritrocitosDokumen9 halamanDeterminación glucosa eritrocitosDiego Sait Cruz HernandezBelum ada peringkat
- M21061B05 Dextrosa 5%Dokumen4 halamanM21061B05 Dextrosa 5%CamilaBelum ada peringkat
- Ciclo de CoriDokumen4 halamanCiclo de CoriJuana Felix MendezBelum ada peringkat
- Metabolismo - Adriana Zubia.3555395Dokumen10 halamanMetabolismo - Adriana Zubia.3555395ADRIANA ZUBIA FLORESBelum ada peringkat
- Coordinación metabólica por el sistema neuroendocrinoDokumen124 halamanCoordinación metabólica por el sistema neuroendocrinoLuna NacusiBelum ada peringkat
- Bioquimica Resumen H 601512 Downloadable 3597272Dokumen14 halamanBioquimica Resumen H 601512 Downloadable 3597272Fabián OlivaresBelum ada peringkat
- Alteraciones Metabólicas PDFDokumen12 halamanAlteraciones Metabólicas PDFSCFranckBelum ada peringkat
- Clase Unica Unidad VIIDokumen53 halamanClase Unica Unidad VIIPedro FernandezBelum ada peringkat
- Practica #11Dokumen14 halamanPractica #11milagros larico100% (1)
- Banco de Preguntas para Final de BioquimicaDokumen47 halamanBanco de Preguntas para Final de Bioquimicavaleria mendezBelum ada peringkat
- Metabolismo de Hidratos de Carbono Tema 9Dokumen37 halamanMetabolismo de Hidratos de Carbono Tema 9Karina BlancoBelum ada peringkat
- CICLO DE CORI Y ALANINA: MECANISMOS DE TRANSPORTE DE ENERGÍA Y NITRÓGENODokumen9 halamanCICLO DE CORI Y ALANINA: MECANISMOS DE TRANSPORTE DE ENERGÍA Y NITRÓGENOEstefania Notario CruzBelum ada peringkat
- La importancia de la glucolisisDokumen45 halamanLa importancia de la glucolisislorena navarreteBelum ada peringkat
- Compilados Choices 188 PagDokumen188 halamanCompilados Choices 188 PagAgustina PinedaBelum ada peringkat
- Integración y Regulación Del MetabolismoDokumen35 halamanIntegración y Regulación Del Metabolismomaria carolina olazabal rodriguezBelum ada peringkat
- Guía de Trabajo Tema 10 y 11Dokumen12 halamanGuía de Trabajo Tema 10 y 11Yazmin DumetBelum ada peringkat
- Conf #7-Gluconeogenesis UASDDokumen28 halamanConf #7-Gluconeogenesis UASDCristhie M. Santana JiménezBelum ada peringkat
- GlucogenólisisDokumen3 halamanGlucogenólisisJosué Delgado Navas100% (1)
- Silabo-Bioquimica IAG 2019-1Dokumen7 halamanSilabo-Bioquimica IAG 2019-1Anonymous ksLPzfCNylBelum ada peringkat
- Gluconeogénesis, vía de las pentosasDokumen17 halamanGluconeogénesis, vía de las pentosasAndrea MoralesBelum ada peringkat
- Resumen metabólico interrelacionesDokumen22 halamanResumen metabólico interrelacionesAngel OnceBelum ada peringkat
- Mitocondrias y Respiración Celularn90Dokumen31 halamanMitocondrias y Respiración Celularn90schneider calderon hidalgoBelum ada peringkat