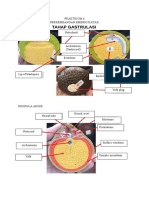
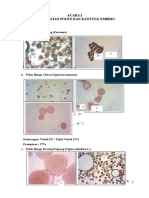
Anda mungkin juga menyukai
- Megasporogenesis Dan MegagametogenesisDokumen8 halamanMegasporogenesis Dan MegagametogenesisiskhawatunamanahBelum ada peringkat
- Embrio SacDokumen12 halamanEmbrio SacicanandaBelum ada peringkat
- Embriogenesis dan Megasporogenesis pada PinaceaeDokumen4 halamanEmbriogenesis dan Megasporogenesis pada PinaceaeLaila Alvi NurinBelum ada peringkat
- OvuleDevelopmentDokumen59 halamanOvuleDevelopmentjensa yuswantoroBelum ada peringkat
- AnthocerosDokumen43 halamanAnthocerosMustika VitaBelum ada peringkat
- Famili TiliaceaeDokumen6 halamanFamili TiliaceaeAtika Ayu KusumaningtyasBelum ada peringkat
- Perkembangan Embrio KatakDokumen9 halamanPerkembangan Embrio KatakAhvaBelum ada peringkat
- Perkembangan Ovulum Markisa (Passiflora SP) PDF PDFDokumen7 halamanPerkembangan Ovulum Markisa (Passiflora SP) PDF PDFicha nissaBelum ada peringkat
- PistilumDokumen23 halamanPistilumFaiza ImawatiBelum ada peringkat
- Organogenesis Pada AvesDokumen7 halamanOrganogenesis Pada AvesRobi CioBelum ada peringkat
- Tugas PoliembrioniDokumen16 halamanTugas PoliembrioniHazan LumzBelum ada peringkat
- LapresDokumen18 halamanLapresLaila Alvi NurinBelum ada peringkat
- Laprak Reproduksi Tumbuha2Dokumen18 halamanLaprak Reproduksi Tumbuha2Kesdik Kusuma Arista Bakti100% (1)
- Perkembangan PteridophytaDokumen4 halamanPerkembangan PteridophytaYusuf BachtiarBelum ada peringkat
- Lapprak Embrio AyamDokumen17 halamanLapprak Embrio AyamNaimmatus SholikhahBelum ada peringkat
- Embriologi AyamDokumen9 halamanEmbriologi AyamSintya Tika DewaBelum ada peringkat
- MikrosporogenesisDokumen54 halamanMikrosporogenesisfaradisa35Belum ada peringkat
- Siklus Hidup PinusDokumen1 halamanSiklus Hidup PinusAdis Indah SariBelum ada peringkat
- PERKEMBANGAN EMBRIO AYAM WHOLEMOUNTDokumen20 halamanPERKEMBANGAN EMBRIO AYAM WHOLEMOUNTMega FrezaaBelum ada peringkat
- Kegiatan 2 Perkembangan Alat Kelamin Bunga MarkisaDokumen41 halamanKegiatan 2 Perkembangan Alat Kelamin Bunga MarkisaDwi rahmaBelum ada peringkat
- Sporogenesis BryophytaDokumen4 halamanSporogenesis BryophytailuBelum ada peringkat
- Kel 4 Embriogenesis BryophytaDokumen20 halamanKel 4 Embriogenesis BryophytaYesi FebriBelum ada peringkat
- (Kegiatan 4) - Tri Purwa Ningrum - 18308141064 - Biologi 2018Dokumen13 halaman(Kegiatan 4) - Tri Purwa Ningrum - 18308141064 - Biologi 2018NingrumBelum ada peringkat
- Makalah Bio RemigesDokumen11 halamanMakalah Bio Remigesartha kurniawanBelum ada peringkat
- GEN LETAL DOMINAN PADA DROSOPHILADokumen37 halamanGEN LETAL DOMINAN PADA DROSOPHILAWahyu TiasBelum ada peringkat
- FILUM ECHINODERMATADokumen3 halamanFILUM ECHINODERMATATjhieahya SyahvillaBelum ada peringkat
- Evolusi dan Sifat MagnoliidDokumen14 halamanEvolusi dan Sifat MagnoliidZaenuddin Ali100% (1)
- Organogenesis Turunan EktodermDokumen38 halamanOrganogenesis Turunan EktodermMuhamad Ikbal Tawakal80% (5)
- Jeno - h1041191066 - Genetika Acara Teknik Pemeriksaan KromosomDokumen12 halamanJeno - h1041191066 - Genetika Acara Teknik Pemeriksaan KromosomJenoBelum ada peringkat
- Evolusi Tumbuhan MonokotilDokumen7 halamanEvolusi Tumbuhan MonokotilPutry Iga UntariBelum ada peringkat
- II B - Amelia Hidayati - 2010119120010 - Laporan Bunga TunggalDokumen27 halamanII B - Amelia Hidayati - 2010119120010 - Laporan Bunga TunggalAmelia HidayatiBelum ada peringkat
- International Code of Nomenclature-2Dokumen25 halamanInternational Code of Nomenclature-2Inaya SetianniBelum ada peringkat
- AvesDokumen13 halamanAvesFajriati RiantobyBelum ada peringkat
- Reproduksi Ovulum Dan Organ Reproduksi BetinaDokumen9 halamanReproduksi Ovulum Dan Organ Reproduksi Betinaneny andriyaniBelum ada peringkat
- DermaDokumen2 halamanDermainayatulBelum ada peringkat
- Pengaruh Lama Penyinaran Sinar Ultraviolet (Uv) Dan Macam Strainterhadap Frekuensi Pindah Silang (Crossing Over)Dokumen30 halamanPengaruh Lama Penyinaran Sinar Ultraviolet (Uv) Dan Macam Strainterhadap Frekuensi Pindah Silang (Crossing Over)Yuswo PurwodarmintoBelum ada peringkat
- MENGUKUR ALAT DAN FUNGSINYADokumen16 halamanMENGUKUR ALAT DAN FUNGSINYASeptianti AmaliaBelum ada peringkat
- Bunga 2013Dokumen29 halamanBunga 2013Nur Rohmah0% (1)
- Neurospora crassa: Jamur Oncom yang Berperan Penting dalam PenelitianDokumen7 halamanNeurospora crassa: Jamur Oncom yang Berperan Penting dalam Penelitianji jiBelum ada peringkat
- Analisis SpermatozoaDokumen13 halamanAnalisis SpermatozoaArfenda Harum LuthfiaBelum ada peringkat
- BUAH DAN BIJINYADokumen5 halamanBUAH DAN BIJINYAnovika dwiBelum ada peringkat
- ANDROESIUMDokumen7 halamanANDROESIUMEs De Pe100% (2)
- Tahap Awal Perkembangan Embrio KatakDokumen18 halamanTahap Awal Perkembangan Embrio KatakFatmawati AtiBelum ada peringkat
- KatakDokumen30 halamanKatakgraciaBelum ada peringkat
- Laporan AnteraDokumen9 halamanLaporan AnteraEmma Maulida100% (1)
- Laporan Praktikum Fabids Kelompok 1Dokumen112 halamanLaporan Praktikum Fabids Kelompok 1sitiBelum ada peringkat
- Pengamatan Polen Dan Kantung Embrio'16Dokumen10 halamanPengamatan Polen Dan Kantung Embrio'16Dessy WulandariBelum ada peringkat
- Angiospermae Dan GymnospermaeDokumen5 halamanAngiospermae Dan GymnospermaeFirman Ali RahmanBelum ada peringkat
- Sporagenesis Dan GametogenesisDokumen17 halamanSporagenesis Dan GametogenesisArlinda Puspita SariBelum ada peringkat
- DAH66533 A 40728 432419026 201027105126 DikonversiDokumen71 halamanDAH66533 A 40728 432419026 201027105126 Dikonversilaila magfirah mahmudBelum ada peringkat
- Kelompok 10 Mikrogametogenesis Dan Mikrosporogenesis Lilium Dan RicinusDokumen24 halamanKelompok 10 Mikrogametogenesis Dan Mikrosporogenesis Lilium Dan RicinusNoorma ParamithaBelum ada peringkat
- PinophytaDokumen29 halamanPinophytaMuhammadIhsanAlawiBelum ada peringkat
- Anatomi BenihDokumen17 halamanAnatomi Benihanon_246834049Belum ada peringkat
- SISTEM SELF INKOMPATIBILITAS TANAMANDokumen4 halamanSISTEM SELF INKOMPATIBILITAS TANAMANauliahasnaBelum ada peringkat
- Embriologi Tumbuhan Jilid II AngiospermaeDokumen36 halamanEmbriologi Tumbuhan Jilid II AngiospermaeMuhammad MahfudhBelum ada peringkat
- EKOMORFOLOGI HEWANDokumen5 halamanEKOMORFOLOGI HEWANRisaul AdikaBelum ada peringkat
- Perkembangan Embryo KatakDokumen18 halamanPerkembangan Embryo KatakFitri SusantiBelum ada peringkat
- MEGASPOROGENESISDokumen2 halamanMEGASPOROGENESISAdri AlfizaBelum ada peringkat
- MATERI Megasporogenesis Dan MegagametogenesisDokumen2 halamanMATERI Megasporogenesis Dan Megagametogenesistashie nadaBelum ada peringkat
- Presentasi SPTDokumen29 halamanPresentasi SPTFebriani DitaBelum ada peringkat