Anda mungkin juga menyukai
- Tercera Semana Embriologia HumanaDokumen33 halamanTercera Semana Embriologia HumanaJhonatan Israel Freire Liquín100% (3)
- Embriologia Del Sistema RespiratorioDokumen44 halamanEmbriologia Del Sistema Respiratorioluisortega93Belum ada peringkat
- Transporte de GametosDokumen12 halamanTransporte de GametosPerla MorenoBelum ada peringkat
- Tercera y Octava Semana de Desarrollo EmbrionarioDokumen6 halamanTercera y Octava Semana de Desarrollo EmbrionarioXiomara LuengoBelum ada peringkat
- Fisiologia FemeninaDokumen3 halamanFisiologia Femeninaastrid viverosBelum ada peringkat
- Cavidades Corporales, Mesenterios y DiafragmaDokumen51 halamanCavidades Corporales, Mesenterios y DiafragmaKaren Hernàndez67% (3)
- GUIA DE EMBRIOLOGIA C ContestadasDokumen3 halamanGUIA DE EMBRIOLOGIA C Contestadasarlet mvBelum ada peringkat
- AnencefaliaDokumen3 halamanAnencefaliaKasamivega100% (1)
- Desarrollo de Cavidades CorporalesDokumen2 halamanDesarrollo de Cavidades CorporalesArantza CuenBelum ada peringkat
- Desarrollo Del Sistema NerviosoDokumen6 halamanDesarrollo Del Sistema NerviosoFelipe Poblete VegaBelum ada peringkat
- Derivados de La Cresta NeuralDokumen4 halamanDerivados de La Cresta NeuralArely Garrido FloresBelum ada peringkat
- Sexta Semana de DesarrolloDokumen52 halamanSexta Semana de DesarrolloGloria Patricia Limon BeilesBelum ada peringkat
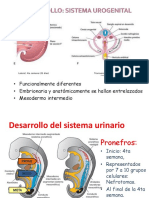
- UrogenitalDokumen40 halamanUrogenitalElianneBelum ada peringkat
- Desarrollo Embrionario de Cabeza y Cuello BlogDokumen37 halamanDesarrollo Embrionario de Cabeza y Cuello BlogpachecquinBelum ada peringkat
- Embarazo MúltipleDokumen7 halamanEmbarazo MúltipleSergio BurgoaBelum ada peringkat
- Moore Embriologia - CAPITULO UNO: Introducción Al Ser Humano en DesarrolloDokumen3 halamanMoore Embriologia - CAPITULO UNO: Introducción Al Ser Humano en DesarrolloSilvia100% (1)
- Dengue y EmbarazoDokumen4 halamanDengue y EmbarazoErick GutierrezBelum ada peringkat
- Embriologia Del Sistema Nervioso DR Carlos Alejandro Gonzales Medina UNMSMDokumen83 halamanEmbriologia Del Sistema Nervioso DR Carlos Alejandro Gonzales Medina UNMSMCarlos GonzalesBelum ada peringkat
- Desarrollo Embriológico Aparato RespiratorioDokumen24 halamanDesarrollo Embriológico Aparato Respiratoriojose eduardo bello ortiz100% (2)
- EmbriologiaDokumen5 halamanEmbriologiaJhey Kenider Benites IzquierdoBelum ada peringkat
- Embriología de Faringe y LaringeDokumen23 halamanEmbriología de Faringe y LaringeChristian Galvan67% (3)
- Embriologia Power Point Ciclo OvaricoDokumen22 halamanEmbriologia Power Point Ciclo OvaricoLivanessa Ubri GonzalezBelum ada peringkat
- Aparato Reproductor FemeninoDokumen13 halamanAparato Reproductor FemeninoCristian Carlos Damian CalleBelum ada peringkat
- Embriología Prac - Ovogenesis - Ciclo MenstrualDokumen37 halamanEmbriología Prac - Ovogenesis - Ciclo MenstrualMiguelIvanB.GuerreroBelum ada peringkat
- Sistema Reproductor FemeninoDokumen78 halamanSistema Reproductor FemeninoIVONXYNBelum ada peringkat
- Angio Genesis y HematogenesisDokumen26 halamanAngio Genesis y HematogenesisMariel Huamancayo100% (1)
- ProtuberanciaDokumen7 halamanProtuberanciaSteven JiménezBelum ada peringkat
- Respiratorio EmbrioDokumen47 halamanRespiratorio EmbrioRosa Angelica Failoc PiscoyaBelum ada peringkat
- Embriología 6Dokumen25 halamanEmbriología 6Nicole DíazBelum ada peringkat
- Embriología CorazónDokumen12 halamanEmbriología CorazónAna SalasBelum ada peringkat
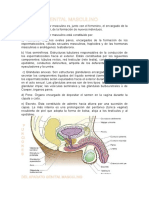
- Embrilogia Del Sistema Genital Masculino y FemenioDokumen27 halamanEmbrilogia Del Sistema Genital Masculino y FemenioOctavio Villacrez OlascoagaBelum ada peringkat
- Desarrollo Embriológico Del Sistema CardiovascularDokumen68 halamanDesarrollo Embriológico Del Sistema CardiovascularYoselin FontBelum ada peringkat
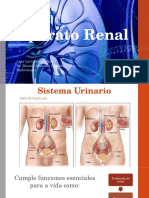
- Desarrollo Del Sistema UrinarioDokumen29 halamanDesarrollo Del Sistema UrinarioAlberto LrBelum ada peringkat
- Aparato Genital Masculino MañanaDokumen9 halamanAparato Genital Masculino MañanaHuamani Flores JhBelum ada peringkat
- Fecundacinsegmentacineimplantacin 121206152556 Phpapp02Dokumen55 halamanFecundacinsegmentacineimplantacin 121206152556 Phpapp02María Esperanza Carrizo GarcíaBelum ada peringkat
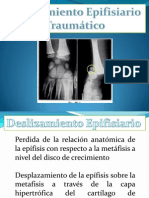
- Deslizamiento Epifisiario TraumaticoDokumen42 halamanDeslizamiento Epifisiario TraumaticoDavid Sztayzel100% (2)
- HISTOLOGÍADokumen55 halamanHISTOLOGÍAConstanza PalavecinoBelum ada peringkat
- Tema 13. Cavidades Corporales y SerosasDokumen8 halamanTema 13. Cavidades Corporales y SerosasVibe NationBelum ada peringkat
- Placenta y Membranas Fetales-Mantilla Ibañez.Dokumen10 halamanPlacenta y Membranas Fetales-Mantilla Ibañez.María Luisa MantillaBelum ada peringkat
- Morfogenesis de La PlacentaDokumen17 halamanMorfogenesis de La PlacentajjgenconBelum ada peringkat
- Fisiologia Gineco MarianDokumen14 halamanFisiologia Gineco Marianjanny Romero100% (1)
- Embarazo MúltipleDokumen9 halamanEmbarazo Múltipleximena vazartBelum ada peringkat
- Gonadas EmbriologiaDokumen1 halamanGonadas EmbriologiaMariana AltamiranoBelum ada peringkat
- Capitulo 55 FisioDokumen2 halamanCapitulo 55 FisioVickyFingadoBelum ada peringkat
- Enfermedad TrofoblasticaDokumen28 halamanEnfermedad TrofoblasticaGuadalupe JojoBelum ada peringkat
- Laboratorio Placenta Embriología UCE 2014Dokumen8 halamanLaboratorio Placenta Embriología UCE 2014John Romero CevallosBelum ada peringkat
- Primera y Segunda Semana Del Desarrollo EmbrionarioDokumen12 halamanPrimera y Segunda Semana Del Desarrollo EmbrionarioFhlip Rams0% (1)
- 3 Ciclo UterinDokumen9 halaman3 Ciclo UterinAnonymous hkjp4qMVRsBelum ada peringkat
- Embriologia Piel Cabeza y CuelloDokumen45 halamanEmbriologia Piel Cabeza y CuelloGabriel Alejandro FloresBelum ada peringkat
- Desarrollo Del Sistema MuscularDokumen31 halamanDesarrollo Del Sistema MuscularMichelle Cartín TristánBelum ada peringkat
- Fisiopatología de CaCuDokumen2 halamanFisiopatología de CaCuArmando D Martínez100% (1)
- Embriologia PreguntasDokumen208 halamanEmbriologia PreguntasCristhian AndréBelum ada peringkat
- Plexocervical 100502194607 Phpapp02Dokumen18 halamanPlexocervical 100502194607 Phpapp02Luis Jesus DkBelum ada peringkat
- Ciclo MenstrualDokumen19 halamanCiclo MenstrualMonica RosarioBelum ada peringkat
- Desarrollo Del Sistema ArterialDokumen25 halamanDesarrollo Del Sistema ArterialAlfonso Beltrán100% (1)
- Embriologia, Anatomia y Fisiologia Del Aparato Reproductor FemeninoDokumen38 halamanEmbriologia, Anatomia y Fisiologia Del Aparato Reproductor FemeninoLitos Montenegro100% (1)
- Semana 11 Práctica EmbriologíaDokumen9 halamanSemana 11 Práctica EmbriologíaAna Quispe IbericoBelum ada peringkat
- Embriologia de EsqueletoDokumen7 halamanEmbriologia de EsqueletoViviana AragonBelum ada peringkat
- Sistema Oseo, HematopoyesisDokumen11 halamanSistema Oseo, HematopoyesisJ. Martín Jilote Mtz.Belum ada peringkat
- Presentació T5 (Ampliado Fotos)Dokumen26 halamanPresentació T5 (Ampliado Fotos)clara100% (1)
- Didactik-Fichas DescriptivasDokumen7 halamanDidactik-Fichas DescriptivasIsaura MaldonadoBelum ada peringkat
- Fallas AceroDokumen33 halamanFallas AceroGustavo Adolfo100% (1)
- Tarea 4Dokumen23 halamanTarea 4fastcarptyBelum ada peringkat
- Tipos de BiopsiasDokumen6 halamanTipos de BiopsiasKevin Carranza VillaoBelum ada peringkat
- Ensayo de Paco YunqueDokumen4 halamanEnsayo de Paco YunqueANTONELLA BELEN RAMOS JARABelum ada peringkat
- Guia 14. Enecuaciones RacionalesDokumen4 halamanGuia 14. Enecuaciones RacionalesAugusto Arturo Querales AmayaBelum ada peringkat
- Unidad 06Dokumen3 halamanUnidad 06Luis Solar Verastegui0% (1)
- TP 6 Salud y Ad 4toDokumen6 halamanTP 6 Salud y Ad 4toMelody AizpitarteBelum ada peringkat
- Franco CiprianiDokumen1 halamanFranco CiprianiAlonso LópezBelum ada peringkat
- Cotizacion Examenes Medicos Agosto AydapackDokumen4 halamanCotizacion Examenes Medicos Agosto AydapackRUBY HBelum ada peringkat
- Los 7 Cielos en La Toráh Con PictográficosDokumen5 halamanLos 7 Cielos en La Toráh Con PictográficosTim Severich100% (3)
- Presentacion Telegram para DocentesDokumen54 halamanPresentacion Telegram para DocentesCarlos Luis Mendoza VelázquezBelum ada peringkat
- Tarea 1 Del Ser Humano y Su Contexto. (Autoguardado)Dokumen7 halamanTarea 1 Del Ser Humano y Su Contexto. (Autoguardado)Manuel Aragonez100% (1)
- RUELAS Maria PEMCDokumen3 halamanRUELAS Maria PEMCMaría Iracema Ruelas AguilarBelum ada peringkat
- Lab1105 4 I 2018Dokumen5 halamanLab1105 4 I 2018Deadpool FulgueraBelum ada peringkat
- 3ero y 4to - Carpeta de Recuperacion MatematicaDokumen29 halaman3ero y 4to - Carpeta de Recuperacion MatematicaAnonymous bDl76usdva0% (1)
- Examen FinalDokumen3 halamanExamen Finalluis angel leon usurinBelum ada peringkat
- Modelo Acta ConstitutivaDokumen9 halamanModelo Acta ConstitutivaBerenice BolañosBelum ada peringkat
- Ledvance® Spotlight Modular LatamDokumen3 halamanLedvance® Spotlight Modular LatamNelydaTaipeBelum ada peringkat
- Ex Fisico RodillaDokumen80 halamanEx Fisico RodillaKawey CaiBelum ada peringkat
- PLANIFICACION DE EdA - 2022Dokumen44 halamanPLANIFICACION DE EdA - 2022David Mendoza GonzalesBelum ada peringkat
- Práctica Estática IIDokumen4 halamanPráctica Estática IIAracely Palomino HuamanBelum ada peringkat
- LIBRE DE ATADURAS ANDERSON NEIL AbusoDokumen194 halamanLIBRE DE ATADURAS ANDERSON NEIL AbusoSilvia Colarte100% (3)
- Revisoria Fiscal Actividad 3Dokumen10 halamanRevisoria Fiscal Actividad 3Lizeth Xiomara RAMIREZ CORREABelum ada peringkat
- Estrategias para El Diagnostico ClinicoDokumen15 halamanEstrategias para El Diagnostico Clinicodebbie38Belum ada peringkat
- Normativa Ecuador Uso GNC en AutomotoresDokumen19 halamanNormativa Ecuador Uso GNC en Automotoresjofre tituañaBelum ada peringkat
- Ejemplo de Aplicación Del Teorema Del Límite Central PDFDokumen1 halamanEjemplo de Aplicación Del Teorema Del Límite Central PDFjwebBelum ada peringkat
- Resumen Termodinamica FundamentalDokumen3 halamanResumen Termodinamica FundamentalMarco Antonio Alonso DelgadilloBelum ada peringkat