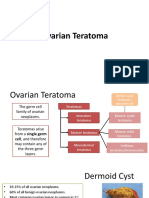
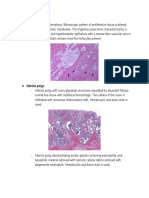
Anda mungkin juga menyukai
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Mol. Cell. Biol.-1999-Di Como-1438-49Dokumen13 halamanMol. Cell. Biol.-1999-Di Como-1438-49Charles J. DiComo, PhDBelum ada peringkat
- Mol. Cell. Biol.-1995-Di Como-1835-46Dokumen12 halamanMol. Cell. Biol.-1995-Di Como-1835-46Charles J. DiComo, PhDBelum ada peringkat
- Exp. Derm.-2008-Di Como-405-11Dokumen7 halamanExp. Derm.-2008-Di Como-405-11Charles J. DiComo, PhDBelum ada peringkat
- Amer. J. Pathol.-2002-Urist-1199-1206Dokumen8 halamanAmer. J. Pathol.-2002-Urist-1199-1206Charles J. DiComo, PhDBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- 2014 Mrcs Osce RECALLDokumen85 halaman2014 Mrcs Osce RECALLIdo EgaziBelum ada peringkat
- Benign Lesions of The OvariesDokumen12 halamanBenign Lesions of The OvariesdocjosmaBelum ada peringkat
- Ovarian Cyst Topic ListDokumen29 halamanOvarian Cyst Topic ListMichelle Defandi ChanBelum ada peringkat
- FGTDokumen248 halamanFGTAmanuel MaruBelum ada peringkat
- Rabinovich 2020Dokumen3 halamanRabinovich 2020Bianca davidBelum ada peringkat
- Chapter 21 Male ReproDokumen34 halamanChapter 21 Male ReproIsfahan MasulotBelum ada peringkat
- Tumor Markers in Surgery: BY DR Joseph AsuquoDokumen22 halamanTumor Markers in Surgery: BY DR Joseph AsuquoIbrahim AkinbolaBelum ada peringkat
- Carcinoma of Prostate: Patho Ogy - Chapter &Dokumen29 halamanCarcinoma of Prostate: Patho Ogy - Chapter &HAMIDBelum ada peringkat
- Testicular TumorsDokumen14 halamanTesticular TumorsJose SirittBelum ada peringkat
- Evaluation of Ovarian Masses in Infants, Children, and Adolescents - UpToDateDokumen22 halamanEvaluation of Ovarian Masses in Infants, Children, and Adolescents - UpToDateScarlette Michelle Jara100% (1)
- Dermoid CystDokumen18 halamanDermoid CystNaila0% (1)
- NeoplasiaDokumen98 halamanNeoplasiaassica0% (1)
- Mature Ovarian Cystic Teratoma Containing Well-Differentiated Cerebellar TissueDokumen2 halamanMature Ovarian Cystic Teratoma Containing Well-Differentiated Cerebellar TissuedrelvBelum ada peringkat
- The 2021 WHO Classification of Tumors of The Central Nervous System, A SumaryDokumen21 halamanThe 2021 WHO Classification of Tumors of The Central Nervous System, A SumaryDTBelum ada peringkat
- 200 Points in Special PathologyDokumen15 halaman200 Points in Special Pathologyjihadeqitaal100% (1)
- Rare Case of Pelvic Schwannoma Mimicking Intra-Ligamenter Uterine Fibroid A Case ReportDokumen5 halamanRare Case of Pelvic Schwannoma Mimicking Intra-Ligamenter Uterine Fibroid A Case ReportSatrya WibawaBelum ada peringkat
- 2017 Immature Ovarian TeratomaDokumen3 halaman2017 Immature Ovarian Teratomapaula rangelBelum ada peringkat
- Understanding Cancer - PPT LectureDokumen169 halamanUnderstanding Cancer - PPT Lecturemirmodepon05100% (15)
- Pathology of Female Genital Tract Short NotesDokumen5 halamanPathology of Female Genital Tract Short NotesameerabestBelum ada peringkat
- Papilloma Benign MelanomaDokumen9 halamanPapilloma Benign MelanomaAdrian John BetanioBelum ada peringkat
- 2 2013 Giant Struma OvariiDokumen2 halaman2 2013 Giant Struma OvariiAnca NeaguBelum ada peringkat
- Benign Gynecologic LesionsDokumen72 halamanBenign Gynecologic LesionsShahbaz KhanBelum ada peringkat
- Neoplastic Diseases of The OvaryDokumen60 halamanNeoplastic Diseases of The OvaryLeslie Ann Pablo MarantanBelum ada peringkat
- Nejmra 2108956Dokumen10 halamanNejmra 2108956silviaBelum ada peringkat
- 8 Diseases of Infancy and ChildhoodDokumen23 halaman8 Diseases of Infancy and ChildhoodBalaji DBelum ada peringkat
- Cleft Palate Caused by Congenital Teratoma: Paediatrics and International Child HealthDokumen5 halamanCleft Palate Caused by Congenital Teratoma: Paediatrics and International Child HealthFitriani MadiBelum ada peringkat
- Histopath ReviewDokumen389 halamanHistopath ReviewLucille MarieBelum ada peringkat
- Nursing OncologyDokumen131 halamanNursing Oncologyapi-3818438100% (5)
- Cchsawa Spring Newsletter Final CompressedDokumen48 halamanCchsawa Spring Newsletter Final Compressedapi-642859049Belum ada peringkat
- Chapter 6 NeoplasiaDokumen24 halamanChapter 6 Neoplasiahenna patelBelum ada peringkat