Anda mungkin juga menyukai
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (73)
- Vox Latina The Pronunciation of Classical LatinDokumen145 halamanVox Latina The Pronunciation of Classical Latinyanmaes100% (4)
- Creature Loot PDF - GM BinderDokumen97 halamanCreature Loot PDF - GM BinderAlec0% (1)
- Chapter 9 Screw ConveyorsDokumen7 halamanChapter 9 Screw ConveyorsMarew Getie100% (1)
- Method Statement Pressure TestingDokumen15 halamanMethod Statement Pressure TestingAkmaldeen AhamedBelum ada peringkat
- Audit AP TestsDokumen3 halamanAudit AP TestsSweet Emme100% (1)
- Cursos Link 2Dokumen3 halamanCursos Link 2Diego Alves100% (7)
- 2016-08-03 Iot Global Forecast Analysis 2015-2025Dokumen65 halaman2016-08-03 Iot Global Forecast Analysis 2015-2025Hoang ThanhhBelum ada peringkat
- Herointroduction 191216 193713 PDFDokumen1 halamanHerointroduction 191216 193713 PDFAnonymous nt8kdDC4RBelum ada peringkat
- Affidavit Responsoring April18Dokumen1 halamanAffidavit Responsoring April18Sreenivas MattaparthiBelum ada peringkat
- ReporterizerDokumen8 halamanReporterizerAnonymous nt8kdDC4RBelum ada peringkat
- Herointroduction 191216 193713Dokumen1 halamanHerointroduction 191216 193713Anonymous nt8kdDC4RBelum ada peringkat
- Training Video Raw MaterialDokumen4 halamanTraining Video Raw MaterialAnonymous nt8kdDC4RBelum ada peringkat
- Img 20180322 0003 PDFDokumen1 halamanImg 20180322 0003 PDFAnonymous nt8kdDC4RBelum ada peringkat
- ReporterizerDokumen8 halamanReporterizerAnonymous nt8kdDC4RBelum ada peringkat
- Img 20180322 0003Dokumen1 halamanImg 20180322 0003Anonymous nt8kdDC4RBelum ada peringkat
- Family Membr Certificts 2d Govt Emplys 2b Issued by Tahasildars Go 145 DT 25-4-15Dokumen2 halamanFamily Membr Certificts 2d Govt Emplys 2b Issued by Tahasildars Go 145 DT 25-4-15api-215249734100% (1)
- Training Video Raw MaterialDokumen4 halamanTraining Video Raw MaterialAnonymous nt8kdDC4RBelum ada peringkat
- ICV document analysis for legal caseDokumen1 halamanICV document analysis for legal caseAnonymous nt8kdDC4RBelum ada peringkat
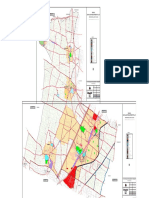
- Mangalagiri Proposed Landuse Map PDFDokumen1 halamanMangalagiri Proposed Landuse Map PDFGurpal kaurBelum ada peringkat
- Kaza Proposed Land Develop MapDokumen1 halamanKaza Proposed Land Develop MapAnonymous nt8kdDC4RBelum ada peringkat
- Draft Perspective Plan for APCRDA Region to 2050Dokumen1 halamanDraft Perspective Plan for APCRDA Region to 2050satishBelum ada peringkat
- Metro Cash Carry India Freedom Sale Print Ad 13th August 2015 VijayawadaDokumen1 halamanMetro Cash Carry India Freedom Sale Print Ad 13th August 2015 VijayawadaAnonymous nt8kdDC4RBelum ada peringkat
- Draft Perspective Plan for APCRDA Region to 2050Dokumen1 halamanDraft Perspective Plan for APCRDA Region to 2050satishBelum ada peringkat
- Girish ResumeDokumen2 halamanGirish ResumeAnonymous nt8kdDC4RBelum ada peringkat
- Draft Detailed Master Plan of Capital City AMARAVATIDokumen1 halamanDraft Detailed Master Plan of Capital City AMARAVATIAnonymous nt8kdDC4RBelum ada peringkat
- Long Standoff Demolition Warheads For Armor, Masonry and Concrete TargetsDokumen27 halamanLong Standoff Demolition Warheads For Armor, Masonry and Concrete Targetsahky7Belum ada peringkat
- Awwa c207 Flanges Spec SheetDokumen13 halamanAwwa c207 Flanges Spec SheetVincent DiepBelum ada peringkat
- Mass ELA Releases ORQs Grade 8Dokumen24 halamanMass ELA Releases ORQs Grade 8scmsliteracyBelum ada peringkat
- Duet Embedded Memories and Logic Libraries For TSMC 28HP: HighlightsDokumen5 halamanDuet Embedded Memories and Logic Libraries For TSMC 28HP: HighlightsmanojkumarBelum ada peringkat
- Commu I ReviewedDokumen45 halamanCommu I ReviewedCabdirisaaq CabdisalanBelum ada peringkat
- Declarative and Procedural Knowledge (Lêda's Final Paper) 2010 01Dokumen13 halamanDeclarative and Procedural Knowledge (Lêda's Final Paper) 2010 01Jair Luiz S. FilhoBelum ada peringkat
- User Manual - Rev3Dokumen31 halamanUser Manual - Rev3SyahdiBelum ada peringkat
- ListDokumen4 halamanListgeralda pierrelusBelum ada peringkat
- New Directions Formative TestDokumen1 halamanNew Directions Formative TestAnonymous xGQcti3qK100% (1)
- 01 Lab ManualDokumen5 halaman01 Lab ManualM Waqar ZahidBelum ada peringkat
- Ferain Et Al, 2016 - The Fatty Acid Profile of Rainbow Trout Liver Cells Modulates Their Tolerance To Methylmercury and CadmiumDokumen12 halamanFerain Et Al, 2016 - The Fatty Acid Profile of Rainbow Trout Liver Cells Modulates Their Tolerance To Methylmercury and Cadmiumarthur5927Belum ada peringkat
- Simple Present 60991Dokumen17 halamanSimple Present 60991Ketua EE 2021 AndrianoBelum ada peringkat
- Regional Office X: Republic of The PhilippinesDokumen2 halamanRegional Office X: Republic of The PhilippinesCoreine Imee ValledorBelum ada peringkat
- Base Is OkDokumen84 halamanBase Is OkajaydevmalikBelum ada peringkat
- Megneto TherapyDokumen15 halamanMegneto TherapyedcanalBelum ada peringkat
- 8510C - 15, - 50, - 100 Piezoresistive Pressure Transducer: Features DescriptionDokumen3 halaman8510C - 15, - 50, - 100 Piezoresistive Pressure Transducer: Features Descriptionedward3600Belum ada peringkat
- PP 12 Maths 2024 2Dokumen21 halamanPP 12 Maths 2024 2Risika SinghBelum ada peringkat
- Chapter - 2 Fish MongeryDokumen10 halamanChapter - 2 Fish MongeryKrishna ChaudharyBelum ada peringkat
- Chenab Railway Bridge Project ReportDokumen50 halamanChenab Railway Bridge Project ReportPreet Chahal100% (1)
- 457 PDFDokumen8 halaman457 PDFAbbey Joy CollanoBelum ada peringkat
- Cambridge IGCSE: Computer Science 0478/12Dokumen16 halamanCambridge IGCSE: Computer Science 0478/12Rodolph Smith100% (2)
- Bond Strength of Normal-to-Lightweight Concrete InterfacesDokumen9 halamanBond Strength of Normal-to-Lightweight Concrete InterfacesStefania RinaldiBelum ada peringkat
- Oxyacetylene Welding (OAW)Dokumen26 halamanOxyacetylene Welding (OAW)athyrahBelum ada peringkat