Anda mungkin juga menyukai
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- PL3257 Tutorial 5 Emotion RegulationDokumen18 halamanPL3257 Tutorial 5 Emotion RegulationRobin TanBelum ada peringkat
- Therapy - Case Study 2Dokumen7 halamanTherapy - Case Study 2api-239581082100% (1)
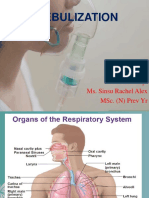
- Nebulization: Presented By: Ms. Sinsu Rachel Alex Msc. (N) Prev YrDokumen29 halamanNebulization: Presented By: Ms. Sinsu Rachel Alex Msc. (N) Prev Yrmanoj reddy100% (1)
- Cold Hip BathDokumen8 halamanCold Hip BathahalyaBelum ada peringkat
- Sixth ScheduleDokumen3 halamanSixth Scheduleangelene koidBelum ada peringkat
- Types of Diagnosis:: Problem-Focused Nursing Diagnoses Have Three Components: (1) Nursing DiagnosisDokumen4 halamanTypes of Diagnosis:: Problem-Focused Nursing Diagnoses Have Three Components: (1) Nursing DiagnosisDaisy MellaBelum ada peringkat
- SBI General Arogya Top Up Policy ProspectusDokumen64 halamanSBI General Arogya Top Up Policy Prospectusyashmpanchal333Belum ada peringkat
- Herpes Simplex Oral: EpidemiologyDokumen5 halamanHerpes Simplex Oral: EpidemiologyFariz RamadhanBelum ada peringkat
- Journal of Clinical Gerontology & Geriatrics: Camilla Jing Hwa Chern, BS, Shyh-Dye Lee, MD, MPHDokumen5 halamanJournal of Clinical Gerontology & Geriatrics: Camilla Jing Hwa Chern, BS, Shyh-Dye Lee, MD, MPHEdith Frederick LiemBelum ada peringkat
- Lactic Acid Bacteria Serum (LABS)Dokumen7 halamanLactic Acid Bacteria Serum (LABS)Felix ValdezBelum ada peringkat
- Three Tiny Little Points On EarDokumen7 halamanThree Tiny Little Points On Eardineshgomber100% (1)
- Millennial Preferences Toward Treating Minor DiseasesDokumen47 halamanMillennial Preferences Toward Treating Minor DiseasesMa Angelica Edison SesucaBelum ada peringkat
- IndianJPain3341-1062069 025700Dokumen10 halamanIndianJPain3341-1062069 025700docbinBelum ada peringkat
- PLR Journey Preview PDFDokumen18 halamanPLR Journey Preview PDFmoumonaBelum ada peringkat
- Measles (Campak, Rubeola, Gabak, Kerumut)Dokumen20 halamanMeasles (Campak, Rubeola, Gabak, Kerumut)Ami UtamiatiBelum ada peringkat
- Pain in Your Butt PreformisDokumen3 halamanPain in Your Butt PreformiscloudoverhillBelum ada peringkat
- 4 Headache PDFDokumen39 halaman4 Headache PDFJohn Ryan ParisBelum ada peringkat
- Emergency Cardiovascular Care Course MatrixDokumen15 halamanEmergency Cardiovascular Care Course MatrixJesus M. Espinosa EchavarriaBelum ada peringkat
- Bad Ragaz Ring MethodDokumen22 halamanBad Ragaz Ring MethodNurdina Afini0% (1)
- DeletedDokumen354 halamanDeletedshalom1413Belum ada peringkat
- Filler Rhinoplasty: Evidence, Outcomes, and ComplicationsDokumen3 halamanFiller Rhinoplasty: Evidence, Outcomes, and ComplicationsАндрей ПетровBelum ada peringkat
- Collaborative Couple Therapy: Page Pr/gurman3.htm&dir Pp/fac&cart - IdDokumen39 halamanCollaborative Couple Therapy: Page Pr/gurman3.htm&dir Pp/fac&cart - IdCarlos InfanteBelum ada peringkat
- Aswathi Haridas 3 YearDokumen24 halamanAswathi Haridas 3 YearLakshmi MuraleedharanBelum ada peringkat
- Running Head: Client Teaching Wound Care 1Dokumen3 halamanRunning Head: Client Teaching Wound Care 1Jerome Lazaro LumanogBelum ada peringkat
- Aerosol TherapyDokumen10 halamanAerosol Therapy04lubna_869632400Belum ada peringkat
- LUTRONIC 2020 CataolgDokumen13 halamanLUTRONIC 2020 Cataolgkhairul ihsanBelum ada peringkat
- Therapeutic Interventions Part I Rubberbands NowDokumen8 halamanTherapeutic Interventions Part I Rubberbands NowAnna Purna100% (1)
- Intraoral Gothic Arch Tracing PDFDokumen5 halamanIntraoral Gothic Arch Tracing PDFSamuel Flores CalderonBelum ada peringkat
- Drying Technology: An International JournalDokumen11 halamanDrying Technology: An International JournalJavier HernandezBelum ada peringkat
- FACTS: There Is No Cure & Complete: Sustained Remissions Are Rare. Principles of TherapyDokumen12 halamanFACTS: There Is No Cure & Complete: Sustained Remissions Are Rare. Principles of TherapyAzizan HannyBelum ada peringkat