Anda mungkin juga menyukai
- Investigación Universitaria Multidisciplinaria 2014Dokumen104 halamanInvestigación Universitaria Multidisciplinaria 2014Investigación Universitaria MultidisciplinariaBelum ada peringkat
- Inv Univ Mult 2013Dokumen200 halamanInv Univ Mult 2013Investigación Universitaria MultidisciplinariaBelum ada peringkat
- Art3 InvUnivMult2013 PDFDokumen8 halamanArt3 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art5 InvUnivMult2013 PDFDokumen9 halamanArt5 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art7 InvUnivMult2013 PDFDokumen8 halamanArt7 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art17 InvUnivMult2013 PDFDokumen9 halamanArt17 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art4 InvUnivMult2013 PDFDokumen9 halamanArt4 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- InvUnivMultB2013 PDFDokumen200 halamanInvUnivMultB2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art1 InvUnivMult2013 PDFDokumen8 halamanArt1 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art2 InvUnivMult2013 PDFDokumen12 halamanArt2 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art6 InvUnivMult2013 PDFDokumen16 halamanArt6 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- GuiaAutoresInvUnivMult2013 PDFDokumen5 halamanGuiaAutoresInvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art15 InvUnivMult2013 PDFDokumen5 halamanArt15 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art16 InvUnivMult2013 PDFDokumen5 halamanArt16 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art19 InvUnivMult2013 PDFDokumen16 halamanArt19 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- AutoresInvUnivMult2013 PDFDokumen8 halamanAutoresInvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art18 InvUnivMult2013 PDFDokumen7 halamanArt18 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- EditorialInvUnivMult2013 PDFDokumen1 halamanEditorialInvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- IndiceInvUnivMult2013 PDFDokumen2 halamanIndiceInvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art14 InvUnivMult2013 PDFDokumen12 halamanArt14 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art11 InvUnivMult2013 PDFDokumen9 halamanArt11 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art13 InvUnivMult2013 PDFDokumen12 halamanArt13 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Inv Univ Mult 2012Dokumen160 halamanInv Univ Mult 2012Investigación Universitaria MultidisciplinariaBelum ada peringkat
- Art12 InvUnivMult2013 PDFDokumen9 halamanArt12 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art9 InvUnivMult2013 PDFDokumen9 halamanArt9 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Editorial InvUnivMult2012Dokumen1 halamanEditorial InvUnivMult2012Investigación Universitaria MultidisciplinariaBelum ada peringkat
- Art8 InvUnivMult2013 PDFDokumen9 halamanArt8 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Art10 InvUnivMult2013 PDFDokumen8 halamanArt10 InvUnivMult2013 PDFInvestigación Universitaria MultidisciplinariaBelum ada peringkat
- Indice InvUnivMult2012Dokumen2 halamanIndice InvUnivMult2012Investigación Universitaria MultidisciplinariaBelum ada peringkat
- GuiaAutores InvUnivMult2012Dokumen5 halamanGuiaAutores InvUnivMult2012Investigación Universitaria MultidisciplinariaBelum ada peringkat
- La Mujer en La Biblia - Opresión y LiberaciónDokumen21 halamanLa Mujer en La Biblia - Opresión y LiberaciónByron Albizures GonzálezBelum ada peringkat
- Resumen 5. Recompensa de La SantidadDokumen1 halamanResumen 5. Recompensa de La SantidadJesusbastos MBelum ada peringkat
- Clase 1 - Sistemas de InyeccionDokumen15 halamanClase 1 - Sistemas de Inyecciondamian floresBelum ada peringkat
- 7 El Pensamiento de PAbloDokumen5 halaman7 El Pensamiento de PAbloAriel CoyosBelum ada peringkat
- Enfoques económicos y relaciones interdisciplinariasDokumen3 halamanEnfoques económicos y relaciones interdisciplinariasGherveth Santis SantizoBelum ada peringkat
- Desarrollo Infantil Primer Año de Vida Interacciones PDFDokumen12 halamanDesarrollo Infantil Primer Año de Vida Interacciones PDFMaría Pastor MayorBelum ada peringkat
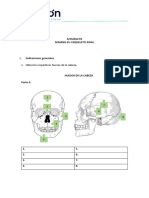
- Tarea Esqueleto AxialDokumen6 halamanTarea Esqueleto AxialdieneBelum ada peringkat
- 6.3.4.4 Packet Tracer - Investigating DUAL FSM InstructionsDokumen4 halaman6.3.4.4 Packet Tracer - Investigating DUAL FSM InstructionsÓscar Vega López100% (1)
- La Evolución y La Sobrepoblación HumanaDokumen1 halamanLa Evolución y La Sobrepoblación HumanaInes LazoBelum ada peringkat
- Informe Práctica 2 EnzimasDokumen3 halamanInforme Práctica 2 EnzimasIsabella RendónBelum ada peringkat
- Clase 1 - Lunes - Construcciones RuralesDokumen74 halamanClase 1 - Lunes - Construcciones RuralesRocio BenítezBelum ada peringkat
- Valvula de Agua BELIMODokumen4 halamanValvula de Agua BELIMOJesús MarquinaBelum ada peringkat
- Dulanto LibroConstrucción SocilaAdolescencia2014 Academia PDFDokumen257 halamanDulanto LibroConstrucción SocilaAdolescencia2014 Academia PDFMaría Estela Fernández100% (1)
- PermanecerEnCristoDokumen91 halamanPermanecerEnCristoEdwin Torres GomezBelum ada peringkat
- Condiciones y Elementos de Las MuestrasDokumen11 halamanCondiciones y Elementos de Las MuestrasMercy Escobar50% (6)
- Edgar Morin Complejidad Restringida y Complejidad GeneralizadaDokumen18 halamanEdgar Morin Complejidad Restringida y Complejidad GeneralizadaNini PoBelum ada peringkat
- Procesos Metalurgicos Laboratorio N 2 inDokumen18 halamanProcesos Metalurgicos Laboratorio N 2 inHeydi CalloBelum ada peringkat
- Ecp Uce 18003 Gcu Id02 0 Gen CD 001 - 0Dokumen55 halamanEcp Uce 18003 Gcu Id02 0 Gen CD 001 - 0David Fernando Bedoya Romero100% (1)
- COMPASS Valvulas Reguladoras CaudalDokumen6 halamanCOMPASS Valvulas Reguladoras Caudaleddy ortegaBelum ada peringkat
- OBJETIVODokumen5 halamanOBJETIVOmarcoBelum ada peringkat
- Antologia de Los Documentos Reales de La Dinastia Carlista - Melchor Ferrer (Org.), 1951Dokumen104 halamanAntologia de Los Documentos Reales de La Dinastia Carlista - Melchor Ferrer (Org.), 1951Eduardo CruzBelum ada peringkat
- Notas para Una Simbología de La RosaDokumen30 halamanNotas para Una Simbología de La Rosaluchinov100% (1)
- Profundiza U5Dokumen14 halamanProfundiza U5Arlex Dauricio Beltran LozanoBelum ada peringkat
- Apunte 11. Dimensionamiento de Pavimentos FlexiblesDokumen264 halamanApunte 11. Dimensionamiento de Pavimentos FlexiblesJhonny Gabriel TorrezBelum ada peringkat
- Fantasma L.J SmithDokumen354 halamanFantasma L.J SmithLina Isabel Lopez Ahumada100% (3)
- Qué Beneficios Podemos Extraer Hoy Del TRABAJO EN REDDokumen5 halamanQué Beneficios Podemos Extraer Hoy Del TRABAJO EN REDcapitan barreraBelum ada peringkat
- Boletín Técnico 12-2022-IPCDokumen16 halamanBoletín Técnico 12-2022-IPCAnthony LunaBelum ada peringkat
- Diagrama de Control para AtributosDokumen23 halamanDiagrama de Control para AtributosGilmarys MartinezBelum ada peringkat
- Clase 33A Excel BASICO - INTERMEDIO - AVANZADO - Ejercicios de Algunos Temas AnterioresDokumen6 halamanClase 33A Excel BASICO - INTERMEDIO - AVANZADO - Ejercicios de Algunos Temas AnterioresLuis PalacioBelum ada peringkat
- Orientación vocacional: elección profesionalDokumen23 halamanOrientación vocacional: elección profesionalLiliana OrtegaBelum ada peringkat