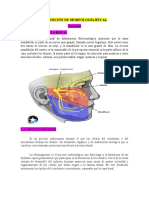
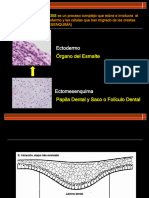
Anda mungkin juga menyukai
- OdontogénesisDokumen10 halamanOdontogénesisRodolfoIgnacioGuerraOsorio100% (1)
- Embriología Del Órgano PulparDokumen24 halamanEmbriología Del Órgano PulparFabiola Lazo CalleBelum ada peringkat
- Desarrollo Dental y Trastornos en Num y FormaDokumen14 halamanDesarrollo Dental y Trastornos en Num y FormaDaniel LoezaBelum ada peringkat
- No 3Dokumen4 halamanNo 3miguelsh0802Belum ada peringkat
- Embriologia Dentaria OdontogenesisDokumen12 halamanEmbriologia Dentaria OdontogenesisDarío Neira100% (1)
- Capítulo 001Dokumen21 halamanCapítulo 001Deceno COLBelum ada peringkat
- ODONTOGENESISDokumen5 halamanODONTOGENESISpopovskiBelum ada peringkat
- ODONTOGENESISDokumen11 halamanODONTOGENESISAndrea QuevedoBelum ada peringkat
- Morfogénesis Del Órgano DentarioDokumen4 halamanMorfogénesis Del Órgano DentarioAripcy MatiasBelum ada peringkat
- Desarrollo de La Denticio Ün2Dokumen17 halamanDesarrollo de La Denticio Ün2José Luis Ardila SalazarBelum ada peringkat
- Histologia BucalDokumen19 halamanHistologia BucalDayana ParragaBelum ada peringkat
- Exposición de Morfología BucalDokumen16 halamanExposición de Morfología BucalHM Wilson AndresBelum ada peringkat
- Morfogénesis Del Órgano DentarioDokumen5 halamanMorfogénesis Del Órgano DentarioOscar Luis100% (1)
- ODONTOGENESISDokumen5 halamanODONTOGENESISbesucona98% (43)
- Resumen 12Dokumen3 halamanResumen 12Diego CastellanosBelum ada peringkat
- Odonto Genesis 3Dokumen72 halamanOdonto Genesis 3Katty OchoaBelum ada peringkat
- 2023 RESUMEN COMPLEJO DENTINOPULPAR Viás de La Pulpa CohenDokumen11 halaman2023 RESUMEN COMPLEJO DENTINOPULPAR Viás de La Pulpa Cohensweetnothingjv0307Belum ada peringkat
- Histología BucodentariaDokumen9 halamanHistología BucodentariaPaloma ScarlethBelum ada peringkat
- Desarrollo y Formación Del Patrón RadicularDokumen5 halamanDesarrollo y Formación Del Patrón RadicularDeiby VacaBelum ada peringkat
- OdontogenesisDokumen60 halamanOdontogenesisNaikaryBelum ada peringkat
- Odontogenesis 2Dokumen54 halamanOdontogenesis 2Sebastián TissoneBelum ada peringkat
- Odontología InfantilDokumen6 halamanOdontología InfantilCalemBelum ada peringkat
- DIENTEDokumen29 halamanDIENTEMaria del Rocio De Lira MonrealBelum ada peringkat
- Embriología DentariaDokumen24 halamanEmbriología DentariaCamila Alexia Quezada ZapataBelum ada peringkat
- Odontogenesis Trabajo ListoDokumen11 halamanOdontogenesis Trabajo ListoCarlos Armando Esqueche AngelesBelum ada peringkat
- Resumen de OdontogénesisDokumen6 halamanResumen de OdontogénesisjuanBelum ada peringkat
- Amelogénesis ImperfectaDokumen5 halamanAmelogénesis Imperfectaalejandrovillalobos2011Belum ada peringkat
- Tarea 1 Origen Embriológico y Constitución de La Pulpa y El PeriodontoDokumen31 halamanTarea 1 Origen Embriológico y Constitución de La Pulpa y El PeriodontoRamírez Martínez Ana PaulinaBelum ada peringkat
- Embriologia Dental ResumenDokumen6 halamanEmbriologia Dental ResumenLuis FerBelum ada peringkat
- Embriología de Cara y CuelloDokumen10 halamanEmbriología de Cara y CuelloTamara AgustoBelum ada peringkat
- OdontogenesisDokumen7 halamanOdontogenesisRicardo Ronny Bejarano LopezBelum ada peringkat
- ODONTOGENESISDokumen61 halamanODONTOGENESISPoligrafia sexto semestreBelum ada peringkat
- Odontoblastos Endondoncia TraducidoDokumen10 halamanOdontoblastos Endondoncia TraducidoArtemetal JaponBelum ada peringkat
- 13 OdontogenesisDokumen30 halaman13 Odontogenesisignacio.odierna.pasqualiniBelum ada peringkat
- Articulo OdontogenesisDokumen20 halamanArticulo OdontogenesisMinerva de SparrowBelum ada peringkat
- OdontopegnesisDokumen20 halamanOdontopegnesisBosco ReyesBelum ada peringkat
- Erupcion Dental y Anomalias DentalesDokumen33 halamanErupcion Dental y Anomalias DentalesulisesmzBelum ada peringkat
- Embriologia Cap 4.Dokumen3 halamanEmbriologia Cap 4.Deyelis W. PerezBelum ada peringkat
- OdontoblastoDokumen6 halamanOdontoblastoJarly Lopez Portocarrero100% (1)
- DENTINOGENESISDokumen12 halamanDENTINOGENESISRUDY ROMARIO BAGUI QUINTEROBelum ada peringkat
- Tema 4 Odontogenesis PDFDokumen68 halamanTema 4 Odontogenesis PDFMIGUEL ANGEL CALLE GRANADABelum ada peringkat
- APOPTOSi SDokumen3 halamanAPOPTOSi SFernanda Comba100% (1)
- Estadio de CampanaDokumen4 halamanEstadio de CampanaPauBelum ada peringkat
- Pulpa DentalDokumen11 halamanPulpa DentalJheremy SolisBelum ada peringkat
- Embriologia de La Cavidad BucalDokumen3 halamanEmbriologia de La Cavidad BucalknowallrpaBelum ada peringkat
- Histologia y Embriologia Bucodental Resumen Completo PDFDokumen26 halamanHistologia y Embriologia Bucodental Resumen Completo PDFana laura felix muñozBelum ada peringkat
- Unidad 2 Anatomía DentalDokumen57 halamanUnidad 2 Anatomía DentalNerea CaracenaBelum ada peringkat
- Filogenia y Ontogenia Del Sistema EstomatognaticoDokumen29 halamanFilogenia y Ontogenia Del Sistema EstomatognaticoStormyBelum ada peringkat
- Embriologia Craneofacial DRA RODRIGUEZDokumen8 halamanEmbriologia Craneofacial DRA RODRIGUEZErika Rodriguez MarteBelum ada peringkat
- Biologia de La PulpaDokumen8 halamanBiologia de La Pulpacarolina herreraBelum ada peringkat
- Odontogenesis 2022-1Dokumen12 halamanOdontogenesis 2022-1valdezbey9Belum ada peringkat
- Embriología de Las Glándulas SalivaresDokumen3 halamanEmbriología de Las Glándulas SalivaresKevin Addler Epquin TafurBelum ada peringkat
- Desarrollo de La LenguaDokumen5 halamanDesarrollo de La LenguaAna Daleska VicuñaBelum ada peringkat
- AmelogenesisDokumen15 halamanAmelogenesisYessie Yance D Aranea100% (3)
- Trabajo de Embriologia OdontogenesisDokumen22 halamanTrabajo de Embriologia OdontogenesisARTBravo20Belum ada peringkat
- Word Traduccion Articulo 1Dokumen12 halamanWord Traduccion Articulo 1Jesus Daniel Segoviano ArmendarizBelum ada peringkat
- Más allá del ADN: La Revolución Epigenética: Desde Mecanismos Celulares hasta Factores Ambientales: Cómo la Epigenética Moldea Nuestro Destino Biológico y las Implicaciones para la Salud, el Comportamiento y el Futuro de la InvestigaciónDari EverandMás allá del ADN: La Revolución Epigenética: Desde Mecanismos Celulares hasta Factores Ambientales: Cómo la Epigenética Moldea Nuestro Destino Biológico y las Implicaciones para la Salud, el Comportamiento y el Futuro de la InvestigaciónBelum ada peringkat
- Conjuntivitis alérgicas: Diagnóstico y tratamientoDari EverandConjuntivitis alérgicas: Diagnóstico y tratamientoBelum ada peringkat
- Caso Clinico Del Servicio de Medicina JaujaDokumen35 halamanCaso Clinico Del Servicio de Medicina JaujaMichael Crisostomo MayhuaBelum ada peringkat
- 8.ejercicio de La Vacuna Antipolio 2013 PreguntasDokumen12 halaman8.ejercicio de La Vacuna Antipolio 2013 PreguntasMichael Crisostomo MayhuaBelum ada peringkat
- 8.ejercicio de La Vacuna Antipolio 2013 PreguntasDokumen12 halaman8.ejercicio de La Vacuna Antipolio 2013 PreguntasMichael Crisostomo MayhuaBelum ada peringkat
- Triptico Imprimir LISTODokumen2 halamanTriptico Imprimir LISTOMichael Crisostomo MayhuaBelum ada peringkat
- Calidad en Los Servicios de SaludDokumen37 halamanCalidad en Los Servicios de SaludMichael Crisostomo Mayhua100% (2)
- Anemia GestacionalDokumen18 halamanAnemia GestacionalMichael Crisostomo MayhuaBelum ada peringkat
- Exposicion Diabetes GestacionalDokumen22 halamanExposicion Diabetes GestacionalMichael Crisostomo Mayhua100% (2)
- Docencia en EnfermeriaDokumen5 halamanDocencia en EnfermeriaMichael Crisostomo Mayhua100% (1)
- Función Investigadora AdministracuionDokumen3 halamanFunción Investigadora AdministracuionMichael Crisostomo Mayhua0% (1)
- Que Es FodaDokumen5 halamanQue Es FodaMichael Crisostomo MayhuaBelum ada peringkat
- Fundamentos de GenéticaDokumen45 halamanFundamentos de GenéticaKevin GutierrezBelum ada peringkat
- Act. Compl. Mat. Gen. Rep. Cel. BDokumen1 halamanAct. Compl. Mat. Gen. Rep. Cel. BSAAVEDRA CANALES VICTORIA ALBERTINABelum ada peringkat
- Guía de Biología IIDokumen4 halamanGuía de Biología IIAbrahan Garcia diazBelum ada peringkat
- Metodo de Reproduccion de La Esponja de MarDokumen3 halamanMetodo de Reproduccion de La Esponja de MarAnderson RamirezBelum ada peringkat
- Cicatriz QueloideDokumen18 halamanCicatriz QueloideFranz RamosBelum ada peringkat
- Infografía Aparato Reproductor Masculino - 20240428 - 192251 - 0000Dokumen3 halamanInfografía Aparato Reproductor Masculino - 20240428 - 192251 - 0000imonio342Belum ada peringkat
- La Clonacion Humana en La ActualidadDokumen2 halamanLa Clonacion Humana en La Actualidadcelso rojas hilarioBelum ada peringkat
- Hickman 2021 18va - Cap 8 Principios Del DesarrolloDokumen29 halamanHickman 2021 18va - Cap 8 Principios Del DesarrolloEverth Anthony Ramos JacintoBelum ada peringkat
- Marco TeóricoDokumen2 halamanMarco TeóricoKatherine A. Mendoza De La CruzBelum ada peringkat
- Unidad1 guiaEstudioLUNESDokumen7 halamanUnidad1 guiaEstudioLUNESStacy SantosBelum ada peringkat
- Primer Parcial 1Q 3ro I y A1Dokumen4 halamanPrimer Parcial 1Q 3ro I y A1George MontañoBelum ada peringkat
- Introducción A La Transcripción Del ADNDokumen97 halamanIntroducción A La Transcripción Del ADNMikado NidomeBelum ada peringkat
- Lisosomas y PeroxisomasDokumen13 halamanLisosomas y Peroxisomaseuclydes BravoBelum ada peringkat
- Biología TP9Dokumen9 halamanBiología TP9Carolina ArriolaBelum ada peringkat
- REGULACIÓNDokumen5 halamanREGULACIÓNCaesarBelum ada peringkat
- Exposicion de EmbriologiaDokumen33 halamanExposicion de EmbriologiaKaren HernandezBelum ada peringkat
- Seminario Ciclo CelularDokumen5 halamanSeminario Ciclo CelularLuis SánchezBelum ada peringkat
- Reproducción en Vertebrados e InvertebradosDokumen2 halamanReproducción en Vertebrados e InvertebradosJanner Melo MoncadaBelum ada peringkat
- Desarrollo Del EspermatozoideDokumen2 halamanDesarrollo Del EspermatozoideJhon Wilner Paredes CondoriBelum ada peringkat
- Cuadernillo de Actividades Biolog-A B-Sica 2023Dokumen70 halamanCuadernillo de Actividades Biolog-A B-Sica 2023lambertoBelum ada peringkat
- Psicología de La Sexualidad (PG 73 85)Dokumen13 halamanPsicología de La Sexualidad (PG 73 85)gloriaBelum ada peringkat
- Biblio Celulas Madre ObtenciónDokumen6 halamanBiblio Celulas Madre ObtenciónEdison Sánchez CepedaBelum ada peringkat
- ORGANISMOSDokumen1 halamanORGANISMOSAileen Kiara Gomez JacintoBelum ada peringkat
- Cien 9 b1 p1 EstDokumen14 halamanCien 9 b1 p1 EstGustavo Cirilo Gazaniga Garrido0% (1)
- Plasticidad NeuronalDokumen14 halamanPlasticidad NeuronalPao RosBelum ada peringkat
- Pnas 2115248119Dokumen7 halamanPnas 2115248119ANA DANIELA FIGUEROA HERNANDEZBelum ada peringkat
- Meiosis Con FormatoDokumen9 halamanMeiosis Con FormatoPaula Andrea Rojas BravoBelum ada peringkat
- Sopa de Letras de Funciones Del Ser HumanoDokumen1 halamanSopa de Letras de Funciones Del Ser HumanoCj QMBelum ada peringkat
- Tema 10. El Núcleo Y La División Celular: Mitosis Y MeiosisDokumen25 halamanTema 10. El Núcleo Y La División Celular: Mitosis Y MeiosisClara Lecanda LorenzoBelum ada peringkat
- Ebau Madrid Biología 2015 Junio Opción ADokumen1 halamanEbau Madrid Biología 2015 Junio Opción APaula Diez OjeaBelum ada peringkat