Anda mungkin juga menyukai
- Counselor Lecturer: Dr. Erna Iftanti, M. PDDokumen15 halamanCounselor Lecturer: Dr. Erna Iftanti, M. PDroat_yeti2255Belum ada peringkat
- ECD Fix.Dokumen46 halamanECD Fix.roat_yeti2255Belum ada peringkat
- Fourth Groups: Example: Jim Read A Book (Jim Is The Subject, Read Is The Action, Book Is The Object)Dokumen2 halamanFourth Groups: Example: Jim Read A Book (Jim Is The Subject, Read Is The Action, Book Is The Object)roat_yeti2255Belum ada peringkat
- Question of The Day 4: 7. I Do Not Know - . - You Are A. When B. Where C. Was D. IfDokumen1 halamanQuestion of The Day 4: 7. I Do Not Know - . - You Are A. When B. Where C. Was D. Ifroat_yeti2255Belum ada peringkat
- Daftar Isi: Happy Children's Day........................ 1Dokumen1 halamanDaftar Isi: Happy Children's Day........................ 1roat_yeti2255Belum ada peringkat
- Table of Content Buku ESPDokumen2 halamanTable of Content Buku ESProat_yeti2255Belum ada peringkat
- Reference: Kelas Semester 1. Tulungagung: CV Sinar Agung AbadiDokumen1 halamanReference: Kelas Semester 1. Tulungagung: CV Sinar Agung Abadiroat_yeti2255Belum ada peringkat
- A Review of Judah Folkman's Remarkable Achievements in BiomedicineDokumen3 halamanA Review of Judah Folkman's Remarkable Achievements in Biomedicineroat_yeti2255Belum ada peringkat
- Hormone Biochemistry, Hormone Biochemistry, Synthesis & Release Synthesis & ReleaseDokumen46 halamanHormone Biochemistry, Hormone Biochemistry, Synthesis & Release Synthesis & Releaseroat_yeti2255Belum ada peringkat
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5795)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1091)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Obstetrics and Gynecology ClinicsDokumen247 halamanObstetrics and Gynecology ClinicsAlexandr TrotskyBelum ada peringkat
- S4 PunctuationDokumen5 halamanS4 PunctuationJake LimBelum ada peringkat
- M Airt TDokumen6 halamanM Airt TLeticia GallardoBelum ada peringkat
- UK Guidelines On The Management of Variceal Hemorrhage in Cirrhotic PatientsDokumen26 halamanUK Guidelines On The Management of Variceal Hemorrhage in Cirrhotic Patientsandus007Belum ada peringkat
- CholestyDokumen17 halamanCholestyapi-3698598Belum ada peringkat
- Welcome Back!: Standard Operating Procedures (SOP) For Resumption of WorkDokumen15 halamanWelcome Back!: Standard Operating Procedures (SOP) For Resumption of WorkSud Joshi100% (1)
- Anticoagulation in EcmoDokumen13 halamanAnticoagulation in Ecmoapi-534214500Belum ada peringkat
- Tropical Health in The Top EndDokumen168 halamanTropical Health in The Top Endemkay1234Belum ada peringkat
- Funda Test 3Dokumen5 halamanFunda Test 3ilove linuxBelum ada peringkat
- Stock Count 01ST July 2021Dokumen4 halamanStock Count 01ST July 2021Williams Lucky ElvisBelum ada peringkat
- Health Week1Dokumen6 halamanHealth Week1Lovely FloresBelum ada peringkat
- Boost Your PA ProtectionDokumen22 halamanBoost Your PA ProtectionLogen TiranBelum ada peringkat
- Anti-Ulcer Activities of Medicinal Plants and Natural ProductsDokumen15 halamanAnti-Ulcer Activities of Medicinal Plants and Natural ProductsMelinda JanuartiBelum ada peringkat
- Case Presentation On LrtiDokumen17 halamanCase Presentation On LrtiNewtan DebBelum ada peringkat
- DHF 2003Dokumen211 halamanDHF 2003Devvi Riasari BunardiBelum ada peringkat
- The Client With Chronic Renal FailureDokumen4 halamanThe Client With Chronic Renal FailureMarisol Jane JomayaBelum ada peringkat
- Intentional Living Worksheet - Copyright - Day Designer - Do Not Distribute - 03-17 PDFDokumen1 halamanIntentional Living Worksheet - Copyright - Day Designer - Do Not Distribute - 03-17 PDFCourtney JohnsonBelum ada peringkat
- NTCC Presentation: Strategic & Competitive Assessment of Brand Looz of Intas PharmmaceuticalsDokumen9 halamanNTCC Presentation: Strategic & Competitive Assessment of Brand Looz of Intas PharmmaceuticalsHarshit VermaBelum ada peringkat
- Dental Academy 2016Dokumen11 halamanDental Academy 2016Priya SargunanBelum ada peringkat
- Tinetti Assessment GaitDokumen1 halamanTinetti Assessment GaitAlexandru ChivaranBelum ada peringkat
- Zollinger Apendicitis PDFDokumen8 halamanZollinger Apendicitis PDFCINDY ADILIA FLORES HERNÁNDEZBelum ada peringkat
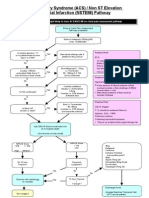
- ACS NSTEMI Clinical PathwayDokumen3 halamanACS NSTEMI Clinical PathwayXtiaRBelum ada peringkat
- Seminar Proposal PTDokumen2 halamanSeminar Proposal PTPeetee MomshieBelum ada peringkat
- HANDOUTDokumen2 halamanHANDOUT27 Nguyễn Thị Huyền TrangBelum ada peringkat
- Case SheetDokumen5 halamanCase SheetvisweswarkcBelum ada peringkat
- Prisma Med - Schneider Electric - or SwitchboardDokumen20 halamanPrisma Med - Schneider Electric - or SwitchboardAna RuxandraBelum ada peringkat
- Mmas4 Dan Mmas8 PDFDokumen1 halamanMmas4 Dan Mmas8 PDFMilly Farisa KurniaBelum ada peringkat
- Rockaway TimesDokumen48 halamanRockaway TimesPeter J. MahonBelum ada peringkat
- Cliff Smyth - Easy Hands and Arms v1-3 - CD Booklet (2004) (Feldenkrais)Dokumen8 halamanCliff Smyth - Easy Hands and Arms v1-3 - CD Booklet (2004) (Feldenkrais)Daived TrombettaBelum ada peringkat
- Epidemiology and Prevention of Vaccine-Preventable Diseases 11th EditionDokumen785 halamanEpidemiology and Prevention of Vaccine-Preventable Diseases 11th EditionitlumBelum ada peringkat