Anda mungkin juga menyukai
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Cap 5 BDokumen15 halamanCap 5 BNina KatiaBelum ada peringkat
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- Cap 4 BDokumen11 halamanCap 4 BNina KatiaBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Capa BDokumen6 halamanCapa BNina KatiaBelum ada peringkat
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- Catherine Cartwright-Jones Henna For HairDokumen60 halamanCatherine Cartwright-Jones Henna For Hairharaputca3Belum ada peringkat
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- CFR 21 Part 117Dokumen43 halamanCFR 21 Part 117almariveraBelum ada peringkat
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Blood Circulation and Transport ModuleDokumen15 halamanBlood Circulation and Transport ModuleIella's Rashid100% (1)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Breakfast - January 2016 PHDokumen84 halamanBreakfast - January 2016 PHAnonymous e2GKJupBelum ada peringkat
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- 2018 Nutritional Monitoring BookDokumen118 halaman2018 Nutritional Monitoring BookrubenBelum ada peringkat
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- Mono Di and MultipleDokumen3 halamanMono Di and MultipleAngel OabelBelum ada peringkat
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- BIOLOGY SS2 3RD TERM E-NOTES (Reviewed)Dokumen89 halamanBIOLOGY SS2 3RD TERM E-NOTES (Reviewed)kanajoseph2009Belum ada peringkat
- Ornamental Flowers and Natatanging FilipinoDokumen10 halamanOrnamental Flowers and Natatanging FilipinoArrelda FuloBelum ada peringkat
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Update Letter PDFDokumen6 halamanUpdate Letter PDFSujeet KumarBelum ada peringkat
- Investigatory ProjectDokumen15 halamanInvestigatory ProjectKailas ArunBelum ada peringkat
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- English Paper 1 SJK Set 2Dokumen15 halamanEnglish Paper 1 SJK Set 2Murely PonnusamyBelum ada peringkat
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (73)
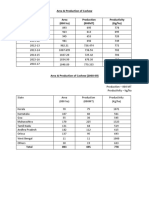
- Area & Production of Cashew Area (000 Ha) Production (000MT) Productivity (Kg/ha)Dokumen6 halamanArea & Production of Cashew Area (000 Ha) Production (000MT) Productivity (Kg/ha)DivyeshBelum ada peringkat
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- Momo Activity 6. Plant Transport Cycle 2Dokumen6 halamanMomo Activity 6. Plant Transport Cycle 2Zherwin Latu-abBelum ada peringkat
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- Wild Edible Fruits of Palpa District, West Nepal: Ras Bihari MahatoDokumen10 halamanWild Edible Fruits of Palpa District, West Nepal: Ras Bihari MahatoAngel Eduardo Moreno AlboresBelum ada peringkat
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Cabra Lunch Menu 5.13.19Dokumen1 halamanCabra Lunch Menu 5.13.19Daniel GerzinaBelum ada peringkat
- Sui Generis Protection of Plant Varieties An Indian PerspectiveDokumen16 halamanSui Generis Protection of Plant Varieties An Indian PerspectiveKarthiayani A.Belum ada peringkat
- Horticulture Development Scenario in Kumaon Region, UttarakhandDokumen10 halamanHorticulture Development Scenario in Kumaon Region, UttarakhandIJAR JOURNALBelum ada peringkat
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- HENGKYDokumen4 halamanHENGKYRahmat HalawaBelum ada peringkat
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- Rubber Wood Processing ManualDokumen80 halamanRubber Wood Processing ManualMustapha GhaniBelum ada peringkat
- DungeonYear A5 Condensed PagesDokumen132 halamanDungeonYear A5 Condensed PagesU- TropuzBelum ada peringkat
- Makers Diet Meals Free Preview ModDokumen40 halamanMakers Diet Meals Free Preview Modolga Penagos100% (6)
- KS TRB LC Read SumDokumen7 halamanKS TRB LC Read Sumgabriela Fernandez MoiBelum ada peringkat
- SOAL PAT Kelas XI SASINGDokumen8 halamanSOAL PAT Kelas XI SASINGsri afrilyaniBelum ada peringkat
- Background of The Study: Citrofortunella Microcarpa (Calamansi) Belongs To The Citrus Family. It Is A FruitDokumen37 halamanBackground of The Study: Citrofortunella Microcarpa (Calamansi) Belongs To The Citrus Family. It Is A FruitTae-tae LachimolalaBelum ada peringkat
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- RPT SC Year 1 (DLP) 2022-2023Dokumen21 halamanRPT SC Year 1 (DLP) 2022-2023norainippgsceBelum ada peringkat
- 1st Grading Summative TestDokumen14 halaman1st Grading Summative TestConnie Diaz CarmonaBelum ada peringkat
- What Are The Differences Between Plants and AnimalsDokumen5 halamanWhat Are The Differences Between Plants and AnimalsWalcot HallBelum ada peringkat
- Kingdom PlantaeDokumen8 halamanKingdom PlantaeDavidBelum ada peringkat
- The Importance of Fasting Saltless and Saltless RecipesDokumen12 halamanThe Importance of Fasting Saltless and Saltless RecipesAnonymous MYpy8rPBelum ada peringkat
- SHS TVL Q1 Module6.SD Present and Store Edited 1Dokumen18 halamanSHS TVL Q1 Module6.SD Present and Store Edited 1Arlene Flor100% (1)
- Tips and Tricks To Crack PMTDokumen3 halamanTips and Tricks To Crack PMTNeyaz HasnainBelum ada peringkat
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)