Anda mungkin juga menyukai
- Fundamental Rights in IndiaDokumen20 halamanFundamental Rights in IndiaRamisha JainBelum ada peringkat
- Maulana Azad National Urdu UniversityDokumen2 halamanMaulana Azad National Urdu UniversityMd QutubuddinBelum ada peringkat
- UGC D.S. Kothari Post-Doc FellowshipsDokumen4 halamanUGC D.S. Kothari Post-Doc FellowshipsalexdruppiBelum ada peringkat
- NPDCL Electrical SyllubusDokumen2 halamanNPDCL Electrical SyllubusMd QutubuddinBelum ada peringkat
- Journal's List - Electrical EngineeringDokumen8 halamanJournal's List - Electrical EngineeringMary MorseBelum ada peringkat
- Electrical Simulation Lab Manual PDFDokumen55 halamanElectrical Simulation Lab Manual PDFAnu tiwari0% (1)
- TeachingDokumen1 halamanTeachingMd QutubuddinBelum ada peringkat
- Retail Supply Tariff Schedule for FY 2018-19Dokumen9 halamanRetail Supply Tariff Schedule for FY 2018-19BabuBelum ada peringkat
- Advertisement PDFDokumen16 halamanAdvertisement PDFPremchand KumarBelum ada peringkat
- Vasa College Recruitment Guide for Electrical EngineeringDokumen5 halamanVasa College Recruitment Guide for Electrical EngineeringjaberyemeniBelum ada peringkat
- 6 ThpayDokumen41 halaman6 Thpaysonaliwani100% (1)
- NotificvhhjationDokumen10 halamanNotificvhhjationsrinureddy217Belum ada peringkat
- Transmission and Distribution Network Reactive PowerDokumen2 halamanTransmission and Distribution Network Reactive PowerMd QutubuddinBelum ada peringkat
- Switched Reluctance Motor ModellingDokumen13 halamanSwitched Reluctance Motor ModellingMd QutubuddinBelum ada peringkat
- Ae Notfn TstranscoDokumen10 halamanAe Notfn TstranscoSuresh DoosaBelum ada peringkat
- What Is GisDokumen64 halamanWhat Is GisMd QutubuddinBelum ada peringkat
- Understanding Permanent Magnet Synchronous MotorsDokumen28 halamanUnderstanding Permanent Magnet Synchronous MotorsMd QutubuddinBelum ada peringkat
- Intorduction To Switched Reluctance MotorDokumen16 halamanIntorduction To Switched Reluctance MotorMd QutubuddinBelum ada peringkat
- Modelling of Power Converter Drives For Induction Motor and PMSMDokumen182 halamanModelling of Power Converter Drives For Induction Motor and PMSMMd QutubuddinBelum ada peringkat
- Modern Control TheoryDokumen1 halamanModern Control TheoryMd QutubuddinBelum ada peringkat
- Introduction To PMSM MachineDokumen5 halamanIntroduction To PMSM MachineMd QutubuddinBelum ada peringkat
- Intorduction To Switched Reluctance MotorDokumen16 halamanIntorduction To Switched Reluctance MotorMd QutubuddinBelum ada peringkat
- Amygdala Ferar BrainDokumen12 halamanAmygdala Ferar BrainMd QutubuddinBelum ada peringkat
- Ultimate Final SCADA PresentationDokumen37 halamanUltimate Final SCADA PresentationJackHarperBelum ada peringkat
- Created by Rick Munz, Edited by Steve Stretz (8/10/2011)Dokumen7 halamanCreated by Rick Munz, Edited by Steve Stretz (8/10/2011)Ashutosh TrivediBelum ada peringkat
- Matlab Code PDFDokumen26 halamanMatlab Code PDFalexwoodwick100% (1)
- Created by Rick Munz, Edited by Steve Stretz (8/10/2011)Dokumen7 halamanCreated by Rick Munz, Edited by Steve Stretz (8/10/2011)Ashutosh TrivediBelum ada peringkat
- Diff. EquationsDokumen12 halamanDiff. EquationsSant NientBelum ada peringkat
- What is GIS? An Introduction to Geographic Information SystemsDokumen36 halamanWhat is GIS? An Introduction to Geographic Information SystemsMd QutubuddinBelum ada peringkat
- Direct Torque Control of Permanent MagnetDokumen56 halamanDirect Torque Control of Permanent MagnetMd QutubuddinBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5784)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (119)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Coordinated Functions of The Nervous, Endocrine, and Reproductive SystemsDokumen30 halamanCoordinated Functions of The Nervous, Endocrine, and Reproductive SystemsSarah CruzBelum ada peringkat
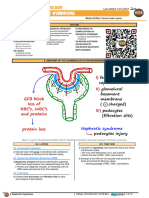
- Renal Pathology - 012) Nephrotic Syndrome (Notes)Dokumen10 halamanRenal Pathology - 012) Nephrotic Syndrome (Notes)hasanatiya41Belum ada peringkat
- Thyroid Disease and Oral Health.Dokumen6 halamanThyroid Disease and Oral Health.crookedgrange6384Belum ada peringkat
- Sensing FeelingDokumen3 halamanSensing FeelingElena UngureanuBelum ada peringkat
- Biliary Microlithiasis, Sludge, Crystals, MicrocrystallizationDokumen6 halamanBiliary Microlithiasis, Sludge, Crystals, MicrocrystallizationniklukBelum ada peringkat
- Male Reproductive System HistologyDokumen12 halamanMale Reproductive System HistologySimbarashe DeshaBelum ada peringkat
- Assessment of Metabolic and Gastrointestinal, Liver AlterationsDokumen10 halamanAssessment of Metabolic and Gastrointestinal, Liver AlterationsGel JamsBelum ada peringkat
- Nervous System Worksheet - Functions and DivisionsDokumen3 halamanNervous System Worksheet - Functions and DivisionsLiezel CauilanBelum ada peringkat
- Electrical conduction system of the heart: an overview of its structure and functionDokumen27 halamanElectrical conduction system of the heart: an overview of its structure and functionAnonymous gUjimJKBelum ada peringkat
- Functional Anatomy of The HeartDokumen44 halamanFunctional Anatomy of The HeartRevanth VejjuBelum ada peringkat
- Z284Dokumen3 halamanZ284Sandeep KaurBelum ada peringkat
- Biology Ch17 - Notes - eDokumen7 halamanBiology Ch17 - Notes - e陳詩淇Belum ada peringkat
- Soal Dan Pembahasan Utbk Inggris 2019 - Rukim - IdDokumen6 halamanSoal Dan Pembahasan Utbk Inggris 2019 - Rukim - IdEkiBheBheBelum ada peringkat
- Thyroid Function TestDokumen2 halamanThyroid Function Testhannah lanasaBelum ada peringkat
- The Endocrine System The Endocrine System: © 2018 Pearson Education, Ltd. 1Dokumen12 halamanThe Endocrine System The Endocrine System: © 2018 Pearson Education, Ltd. 1lourd nabBelum ada peringkat
- Year One Anatomy and Physiology Study NotesDokumen94 halamanYear One Anatomy and Physiology Study Notescoolpinkgurl100% (8)
- Case Study CholelithiasisDokumen14 halamanCase Study Cholelithiasisb_faye20Belum ada peringkat
- NCM 112 Pre Lim NotesDokumen28 halamanNCM 112 Pre Lim NotesYancy TanBelum ada peringkat
- HY Immunology by Mehlaman - PDF Version 1Dokumen41 halamanHY Immunology by Mehlaman - PDF Version 1Drekola KhanBelum ada peringkat
- The Human HeartDokumen1 halamanThe Human HeartMae TanBelum ada peringkat
- Respiratory System: A) The Conducting Portion, Which Consists of The Nasal CavitiesDokumen7 halamanRespiratory System: A) The Conducting Portion, Which Consists of The Nasal CavitiesSalmah Alimah FitriBelum ada peringkat
- What Is Anatomy-WPS OfficeDokumen4 halamanWhat Is Anatomy-WPS OfficePairene TalibBelum ada peringkat
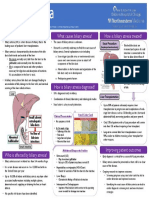
- Chronic Kidney Disease: Causes, Symptoms, Diagnosis and TreatmentDokumen67 halamanChronic Kidney Disease: Causes, Symptoms, Diagnosis and TreatmentpashaBelum ada peringkat
- Homeostasis EssayDokumen2 halamanHomeostasis EssayMysterie_08Belum ada peringkat
- Final Compre Exam Key AnswersDokumen14 halamanFinal Compre Exam Key Answersmj CanilangBelum ada peringkat
- Anatomy MusclesDokumen76 halamanAnatomy MusclesMathios TigerosBelum ada peringkat
- (Hepatoportoenterostomy) : Anna M. Banc-Husu, MDDokumen1 halaman(Hepatoportoenterostomy) : Anna M. Banc-Husu, MDwatimelawatiBelum ada peringkat
- Hypothyrodism in PregnancyDokumen32 halamanHypothyrodism in Pregnancymohammed makki0% (1)
- Insider Guide 11Dokumen144 halamanInsider Guide 11sam100% (3)
- Netter Atlas of Human Neuroscience PDFDokumen310 halamanNetter Atlas of Human Neuroscience PDFAndra Iordache67% (3)