Anda mungkin juga menyukai
- MeningitisBacterianaSintomasDiagnosticoDokumen11 halamanMeningitisBacterianaSintomasDiagnosticoEmily MariñosBelum ada peringkat
- Intoxicacion Por Raticidas AnticoagulantesDokumen20 halamanIntoxicacion Por Raticidas AnticoagulantesMaria Paulina Bueno CuervoBelum ada peringkat
- Cardiopatías CongénitasDokumen52 halamanCardiopatías CongénitasMaria Paulina Bueno CuervoBelum ada peringkat
- Convulsiones febriles: guía para padresDokumen10 halamanConvulsiones febriles: guía para padresMaria Paulina Bueno CuervoBelum ada peringkat
- 1 Electrocardiograma PDFDokumen11 halaman1 Electrocardiograma PDFKely Jhovana Niño SotoBelum ada peringkat
- Activación y Función de Células T y BDokumen9 halamanActivación y Función de Células T y BMaria Paulina Bueno CuervoBelum ada peringkat
- Manual CTO 6ed - FisiologíaDokumen37 halamanManual CTO 6ed - Fisiologíameme100% (21)
- Monografia. Historia Clinica PediatricaDokumen41 halamanMonografia. Historia Clinica PediatricaBetsy Tipacti100% (1)
- Encuesta de SaludDokumen1 halamanEncuesta de SaludAnonymous BFM6zwHBelum ada peringkat
- Preparación de MedicamentosDokumen12 halamanPreparación de Medicamentosshanny barraza100% (2)
- G8 - Dermatosis InflamatoriasDokumen4 halamanG8 - Dermatosis InflamatoriasluzmargaritajcmailcomBelum ada peringkat
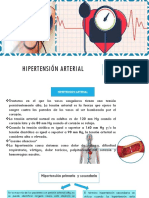
- Hipertension ArterialDokumen41 halamanHipertension ArterialEdy Pol Quispitupa DegadoBelum ada peringkat
- Valoracion de Pruebas Diagnosticas y de Cribaje EXPODokumen26 halamanValoracion de Pruebas Diagnosticas y de Cribaje EXPOPaola Heredia SotoBelum ada peringkat
- PartoDokumen36 halamanPartoSofia AmuchasteguiBelum ada peringkat
- Cuidados Paliativos OpsDokumen5 halamanCuidados Paliativos OpsAndy AvesBelum ada peringkat
- Norma 168 Presentacion FinalDokumen19 halamanNorma 168 Presentacion Finalmiki_mouse_28Belum ada peringkat
- Enfermedad Tromboembolica Venosa BaltazaresDokumen525 halamanEnfermedad Tromboembolica Venosa BaltazaresJulianoEscudeiro100% (1)
- Ritmos cardíacos desfibrilables y no desfibrilables RCPDokumen10 halamanRitmos cardíacos desfibrilables y no desfibrilables RCPSuzy Kiara Park100% (1)
- Bioquímica clínica enzimasDokumen40 halamanBioquímica clínica enzimaszenia garciaBelum ada peringkat
- Exudado FaringeoDokumen8 halamanExudado FaringeoQUIMICO CLINICO WILLIANS SANCHEZ100% (4)
- Conceptos y Principios Generales de InmunizaciónDokumen25 halamanConceptos y Principios Generales de InmunizaciónIldefonso SanchezBelum ada peringkat
- Terapia de Danza para AlzheimerDokumen45 halamanTerapia de Danza para AlzheimerLaura ApaBelum ada peringkat
- Estimulantes Del SNCDokumen7 halamanEstimulantes Del SNCDANIEL ALEJANDROBelum ada peringkat
- Parcial NeuropsicologiaDokumen5 halamanParcial NeuropsicologiaNidissx Bayona0% (1)
- Semiologia EndocrinoDokumen4 halamanSemiologia EndocrinoMaricieloBelum ada peringkat
- Anamnesis Tipo Motricidad OrofacialDokumen2 halamanAnamnesis Tipo Motricidad Orofacialxyd7qmwqznBelum ada peringkat
- Apunte ObstetriciaDokumen121 halamanApunte ObstetriciaJulietaS.LopezNanniniBelum ada peringkat
- Clase 9 Oxigenoterapia, Alimentación Asistida y Cuidados PediátricosDokumen36 halamanClase 9 Oxigenoterapia, Alimentación Asistida y Cuidados PediátricosJose Luis Del RioBelum ada peringkat
- s6 PPT ANTIMICROBIANOSDokumen30 halamans6 PPT ANTIMICROBIANOSLizbeth Honores VasquezBelum ada peringkat
- Planificación Familiar para Quinto Grado de SecundariaDokumen4 halamanPlanificación Familiar para Quinto Grado de SecundariaFlorecita Barrena fustamanteBelum ada peringkat
- Cirrosis hepática y alteraciones bioquímicasDokumen6 halamanCirrosis hepática y alteraciones bioquímicasErika CordovaBelum ada peringkat
- EXPOCIRUGIADokumen19 halamanEXPOCIRUGIAEli Daniel V. AldazBelum ada peringkat
- Cuestionarios-Ipaq-Parq-Anamensis (1) Diego CelisDokumen6 halamanCuestionarios-Ipaq-Parq-Anamensis (1) Diego CelisDiego Fernando Celis LozanoBelum ada peringkat
- 6-Modelo de Examen Final Clinica 2 MICDokumen6 halaman6-Modelo de Examen Final Clinica 2 MICabdallah hamdan60% (5)
- Examen parcial de Medicina del TrabajoDokumen14 halamanExamen parcial de Medicina del TrabajoFERNEYBelum ada peringkat
- Ectasias CornealesDokumen50 halamanEctasias CornealesEmmanuel Aguilar100% (3)
- Directorio Nacional Establecimientos Residenciales CONADICDokumen292 halamanDirectorio Nacional Establecimientos Residenciales CONADICMaria del Carmen Guillén AlcarazBelum ada peringkat