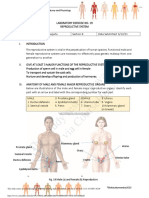
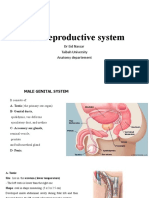
Anda mungkin juga menyukai
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5782)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (587)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (72)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (265)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (119)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Kami Export - Imman Sheikh - Sexual and Asexual HandoutDokumen3 halamanKami Export - Imman Sheikh - Sexual and Asexual HandoutImman SheikhBelum ada peringkat
- Placenta PreviaDokumen4 halamanPlacenta PreviaJeizel IgnacioBelum ada peringkat
- 8 Science NCERT Solutions Chapter 10Dokumen7 halaman8 Science NCERT Solutions Chapter 10Raj Anand100% (1)
- Chapter 3Dokumen6 halamanChapter 3MoiMoi P.S.Belum ada peringkat
- Week 1 What Is Biology Characteristics of LifeDokumen16 halamanWeek 1 What Is Biology Characteristics of LifeAsrar MohamedBelum ada peringkat
- Durian Grafting TechniquesDokumen16 halamanDurian Grafting Techniquesatoydequit100% (1)
- Science 5 Process of FertilizationDokumen4 halamanScience 5 Process of FertilizationMayden GubotBelum ada peringkat
- EDCS 640-Placed-Based Lesson-Plan Marie Pulu ExampleDokumen6 halamanEDCS 640-Placed-Based Lesson-Plan Marie Pulu ExampleMakanani AuauBelum ada peringkat
- Must-Know Key Concepts in PSLE ScienceDokumen30 halamanMust-Know Key Concepts in PSLE ScienceClarisa CynthiaBelum ada peringkat
- Cambridge IGCSE: BIOLOGY 0610/31Dokumen20 halamanCambridge IGCSE: BIOLOGY 0610/31Balachandran PalaniandyBelum ada peringkat
- Reproductive Anatomy: Chapter 34 Lesson 1 Page 958Dokumen13 halamanReproductive Anatomy: Chapter 34 Lesson 1 Page 958JanaBelum ada peringkat
- Endocrinology of Genitourinary SystemDokumen4 halamanEndocrinology of Genitourinary SystemGerardLumBelum ada peringkat
- Laboratory 1 Seed MorphologicalDokumen17 halamanLaboratory 1 Seed MorphologicalJohn Rey De Los SantosBelum ada peringkat
- ScienxeDokumen35 halamanScienxeBloom rachBelum ada peringkat
- Science 10 - Reproductive SystemDokumen32 halamanScience 10 - Reproductive SystemHanabi Scarlet ShadowBelum ada peringkat
- 8- General embryolog د.خالد ميلاد (@USEM - BOT)Dokumen28 halaman8- General embryolog د.خالد ميلاد (@USEM - BOT)Ayoub ZaatoutBelum ada peringkat
- Lab Exercise19 Reproductive System PDFDokumen3 halamanLab Exercise19 Reproductive System PDFcsolutionBelum ada peringkat
- GAMETOGENESIS - SpermatogenisisDokumen40 halamanGAMETOGENESIS - Spermatogenisissteam inventoryBelum ada peringkat
- A History of Developments To Improve in Vitro Ferti Lizati OnDokumen4 halamanA History of Developments To Improve in Vitro Ferti Lizati OnichaBelum ada peringkat
- BO-09 Taxanomy and Embryology of Angiosperms Section - ADokumen12 halamanBO-09 Taxanomy and Embryology of Angiosperms Section - ATanishq PrajapatBelum ada peringkat
- Molar Pregnancy - Case PresentationDokumen7 halamanMolar Pregnancy - Case Presentationアンナドミニク100% (1)
- Ans BOT CH-7 Gymnosperms MCQ TESTDokumen2 halamanAns BOT CH-7 Gymnosperms MCQ TESTIshrak ZamanBelum ada peringkat
- Text Test Bank 11Dokumen11 halamanText Test Bank 11n1n2n3nBelum ada peringkat
- And Organogenesis: in Vitro Culture of Safflower L. Cv. Bhima: Initiation, Growth OptimizationDokumen2 halamanAnd Organogenesis: in Vitro Culture of Safflower L. Cv. Bhima: Initiation, Growth OptimizationAlexandra GalanBelum ada peringkat
- Cross Breeding Systems For Beef CattleDokumen8 halamanCross Breeding Systems For Beef CattleLivestock Managementist IubBelum ada peringkat
- BiologyDokumen10 halamanBiologyNur FarhanimBelum ada peringkat
- Floral Formula: BotanyDokumen7 halamanFloral Formula: BotanyzedoksBelum ada peringkat
- Protimo Antimicrobial Agent Introduction-2013-08-08 PDFDokumen27 halamanProtimo Antimicrobial Agent Introduction-2013-08-08 PDF連宗興Belum ada peringkat
- Anatomy and PhysiologyDokumen2 halamanAnatomy and PhysiologyanglogaliboBelum ada peringkat
- Male Reproductive System MLTDokumen12 halamanMale Reproductive System MLTAbdulrahman MohammedBelum ada peringkat