Anda mungkin juga menyukai
- Três Agulhas e Auriculoterapia para Epicondilite Lateral Um Estudo de CasoDokumen39 halamanTrês Agulhas e Auriculoterapia para Epicondilite Lateral Um Estudo de CasoGláucia LunaBelum ada peringkat
- Apostila Doenças Da TireoideDokumen3 halamanApostila Doenças Da TireoideRita Ferreira100% (1)
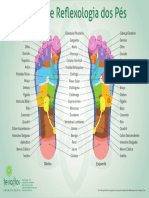
- Mapa de Re Exologia Dos Pés: DireitoDokumen1 halamanMapa de Re Exologia Dos Pés: DireitoGláucia LunaBelum ada peringkat
- Laserpuntura 1 PDFDokumen90 halamanLaserpuntura 1 PDFRogéria Cavalcante Proença100% (1)
- Ícones dos atributos de qualificação dos prestadoresDokumen78 halamanÍcones dos atributos de qualificação dos prestadoresGláucia LunaBelum ada peringkat
- Memento Fitoterápico Da Farmacopéia Brasileira PDFDokumen115 halamanMemento Fitoterápico Da Farmacopéia Brasileira PDFAroldo J. de OliveiraBelum ada peringkat
- Função Energética Dos Pontos: Objeto de Aprendizagem Desenvolvido Por Dr. Ephraim Ferreira MedeirosDokumen57 halamanFunção Energética Dos Pontos: Objeto de Aprendizagem Desenvolvido Por Dr. Ephraim Ferreira MedeirosGláucia LunaBelum ada peringkat
- Gustavo Forlani SoaresDokumen93 halamanGustavo Forlani SoaresGláucia LunaBelum ada peringkat
- SangriaDokumen23 halamanSangriaGláucia LunaBelum ada peringkat
- Afecções NeurologicasDokumen67 halamanAfecções NeurologicasMega TevimBelum ada peringkat
- Códigos de Grabovoi para cura e evoluçãoDokumen101 halamanCódigos de Grabovoi para cura e evoluçãolarie fernandes93% (14)
- !2lcupuntura: E A Mol (AbustifuDokumen143 halaman!2lcupuntura: E A Mol (AbustifuGláucia LunaBelum ada peringkat
- ReflexologiaDokumen87 halamanReflexologiaBianca SilvaBelum ada peringkat
- Formula Rio de Fitoterapicos Da Farmacopeia BrasileiraDokumen126 halamanFormula Rio de Fitoterapicos Da Farmacopeia BrasileirahpascoaBelum ada peringkat
- Mao Pe Reflexologia 2018 PDFDokumen89 halamanMao Pe Reflexologia 2018 PDFRenato Paredes AraújoBelum ada peringkat
- 1 4960792655442215010Dokumen35 halaman1 4960792655442215010Gláucia LunaBelum ada peringkat
- Tung 2013 SPDokumen9 halamanTung 2013 SPMarcos Martini100% (3)
- 51 - O Uso Da Acupuntura Japonesa No Tratamento de CrianYasDokumen21 halaman51 - O Uso Da Acupuntura Japonesa No Tratamento de CrianYasLeonardo Dos Santos SilvaBelum ada peringkat
- Mapa de Re Exologia Dos Pés: DireitoDokumen1 halamanMapa de Re Exologia Dos Pés: DireitoGláucia LunaBelum ada peringkat
- Função Energética Dos Pontos: Objeto de Aprendizagem Desenvolvido Por Dr. Ephraim Ferreira MedeirosDokumen57 halamanFunção Energética Dos Pontos: Objeto de Aprendizagem Desenvolvido Por Dr. Ephraim Ferreira MedeirosGláucia LunaBelum ada peringkat
- Ficha - Leitura Do PulsoDokumen4 halamanFicha - Leitura Do PulsoGláucia LunaBelum ada peringkat
- Afecções NeurologicasDokumen67 halamanAfecções NeurologicasMega TevimBelum ada peringkat
- Livro de EsparadrapoterapiaDokumen45 halamanLivro de Esparadrapoterapiaamfxx89% (28)
- Ajuda AnamneseDokumen7 halamanAjuda AnamneseGláucia LunaBelum ada peringkat
- Mapas Auriculoterapia PDFDokumen15 halamanMapas Auriculoterapia PDFTamara Velloso100% (7)
- Ficha de AnamneseDokumen16 halamanFicha de AnamneseGláucia LunaBelum ada peringkat
- Teorias BasicasDokumen65 halamanTeorias BasicasLanny50000% (1)
- Acupuntura CRFDokumen44 halamanAcupuntura CRFrenatafj100% (1)
- Técnica Do Esparadrapo - Abigail Muniz CaracikiDokumen21 halamanTécnica Do Esparadrapo - Abigail Muniz CaracikiPatricia Pappacena100% (1)
- Ventosa Terapia: O Resgate da Antiga Arte da LongevidadeDokumen2 halamanVentosa Terapia: O Resgate da Antiga Arte da LongevidadeMarcos LobatoBelum ada peringkat
- Densidade de materiaisDokumen2 halamanDensidade de materiaisPaulo Viegas0% (1)
- Orgânica3 Hidrocarbonetos (Parte1)Dokumen21 halamanOrgânica3 Hidrocarbonetos (Parte1)Lorenzo RamosBelum ada peringkat
- Estatuto do servidor DiademaDokumen46 halamanEstatuto do servidor Diademajmaj jmajBelum ada peringkat
- Aposto e VocativoDokumen3 halamanAposto e VocativoJessica VasconcelosBelum ada peringkat
- Engenharia e inovação: compartilhando experiênciasDokumen40 halamanEngenharia e inovação: compartilhando experiênciasFabiano AlvesBelum ada peringkat
- Abordagem metodológica para problemas complexos em planos diretores hospitalaresDokumen40 halamanAbordagem metodológica para problemas complexos em planos diretores hospitalaresJeffrey HaithBelum ada peringkat
- CatalogoDokumen72 halamanCatalogoQue torcida é essa.Belum ada peringkat
- Trading in Zone - Portugues - PDF - Análise Técnica - LuaDokumen184 halamanTrading in Zone - Portugues - PDF - Análise Técnica - LuaRudy Gay StruamBelum ada peringkat
- Livro Natureza RenascimentoDokumen2 halamanLivro Natureza RenascimentoCarolina SenraBelum ada peringkat
- Inquerito - PFDokumen10 halamanInquerito - PFliscappiBelum ada peringkat
- Mapa - Far - Farmacologia Aplicada - 52-2023Dokumen2 halamanMapa - Far - Farmacologia Aplicada - 52-2023Cavalini Assessoria AcadêmicaBelum ada peringkat
- Morte e Vida Severina de João CabralDokumen28 halamanMorte e Vida Severina de João CabralAndré MoreiraBelum ada peringkat
- Halliday 3 - Cap 31Dokumen8 halamanHalliday 3 - Cap 31Peter DutaBelum ada peringkat
- Conceitualização Cognitiva: Pensamentos, Crenças e EstratégiasDokumen5 halamanConceitualização Cognitiva: Pensamentos, Crenças e EstratégiasMauricio Marane0% (1)
- Fibra Tipo 3Dokumen5 halamanFibra Tipo 3Edimilson LanaBelum ada peringkat
- Growth suplementos pagamentoDokumen1 halamanGrowth suplementos pagamentoEu sou o pai do YOUTUBEBelum ada peringkat
- Manual do Aluno Village Park DogDokumen4 halamanManual do Aluno Village Park DogSadm SorocabaBelum ada peringkat
- 2021 BMW 3 Series 28Dokumen474 halaman2021 BMW 3 Series 28Thiago MolverstetBelum ada peringkat
- Procedimentos para ensaio de viga de concreto protendidaDokumen66 halamanProcedimentos para ensaio de viga de concreto protendidaLucasBelum ada peringkat
- Síndrome do chicote: revisão sobre lesão por aceleração cervicalDokumen5 halamanSíndrome do chicote: revisão sobre lesão por aceleração cervicalCatarina DiasBelum ada peringkat
- Análise de Risco para Supressão VegetalDokumen12 halamanAnálise de Risco para Supressão VegetalAdilson Sousa LimaBelum ada peringkat
- Ap3 Pesquisa Mercadologica Uninta - Passei DiretoDokumen11 halamanAp3 Pesquisa Mercadologica Uninta - Passei DiretoFelipe LimaBelum ada peringkat
- 2º Encontro Ciências 15-03-2022Dokumen14 halaman2º Encontro Ciências 15-03-2022Edson AraujoBelum ada peringkat
- Características à vazio e de curto-circuito de máquinas síncronasDokumen6 halamanCaracterísticas à vazio e de curto-circuito de máquinas síncronasCláudia FonsecaBelum ada peringkat
- Apostila 02 - Educação Física-EJA-2º Bimestre - 2ºquadrimestre - FASE IXDokumen2 halamanApostila 02 - Educação Física-EJA-2º Bimestre - 2ºquadrimestre - FASE IXBruna AuadBelum ada peringkat
- A Vida de Uma Princesa Árabe Revela a Opressão das MulheresDokumen144 halamanA Vida de Uma Princesa Árabe Revela a Opressão das MulheresMarcos Bezerra da CostaBelum ada peringkat
- CO MUELLER - Bomba KSB 050-032-250 - Folha de DadosDokumen6 halamanCO MUELLER - Bomba KSB 050-032-250 - Folha de DadosmarceloBelum ada peringkat
- A AMIZADE É MAGICA MODULO OP VFINAL - Por Onze#6904Dokumen27 halamanA AMIZADE É MAGICA MODULO OP VFINAL - Por Onze#6904Thecla CoronataBelum ada peringkat
- Regulamenta acautelamento de material bélico SEAPDokumen26 halamanRegulamenta acautelamento de material bélico SEAPfaelcassioBelum ada peringkat