44 Mecanismos Generales de Protección
Diunggah oleh
Adriana OlivaresJudul Asli
Hak Cipta
Format Tersedia
Bagikan dokumen Ini
Apakah menurut Anda dokumen ini bermanfaat?
Apakah konten ini tidak pantas?
Laporkan Dokumen IniHak Cipta:
Format Tersedia
44 Mecanismos Generales de Protección
Diunggah oleh
Adriana OlivaresHak Cipta:
Format Tersedia
Mecanismos protectores generales
Conceptos bsicos sobre el sistema inmunitario El trmino inmunidad tiene su origen en un vocablo romano que significa privilegio de exencin o estar libre y que hace referencia a la capacidad que poseen los seres vivos de no sufrir continuamente las enfermedades que ocasionan la agresin de los microorganismos. El sistema inmunitario (SI) protege al organismo de una amplia variedad de agentes infecciosos (bacterias, hongos, parsitos y virus) que pueden ocasionar en el organismo que los recibe diferentes enfermedades. Para ello es capaz de reconocer a los componentes del agente patgeno e iniciar una serie de respuestas encaminadas a eliminarlo cuyas caractersticas fundamentales son la especificidad y la memoria. Desde una concepcin clsica se ha hablado de inmunidad humoral cuando la respuesta inmunitaria est mediada por anticuerpos y de inmunidad celular cuando est mediada por clulas. Ambos tipos de respuesta pueden tener la caracterstica de ser especficas a un determinado patgeno o por el contrario producirse de un modo general e inespecfico; mientras que los anticuerpos son los efectores especficos de la inmunidad humoral, el complemento y algunos interferones lo son inespecficos; mientras que los linfocitos T citotxicos son los efectores especficos de la inmunidad celular, la fagocitosis, las clulas asesinas naturales, la citotoxicidad dependiente de anticuerpos y los mediadores solubles son efectores inespecficos de la inmunidad celular. Las principales clulas que participan en las respuestas inmunitarias son los leucocitos, los glbulos blancos de la sangre, de los que se distinguen varios tipos siendo los principales los linfocitos y los fagocitos que, mediante su presencia y la secrecin de diferentes sustancias solubles que son capaces de producir, median en la respuesta del SI ante una agresin. Las disfunciones del SI se pueden entender en una triple vertiente: Respuesta inmunitaria exagerada (hipersensibilidad), respuesta inmunitaria ineficaz (inmunodeficiencia) y reaccin inadecuada frente a autoantigenos (enfermedad autoinmune). Concepto de antgeno y de anticuerpo. Se entiende como antgeno (Ag) cualquier molcula que puede ser reconocida especficamente por cualquiera de los componentes del SI; en un sentido ms restrictivo se entiende como Ag cualquier molcula capaz de inducir la produccin de anticuerpos especficos. Los anticuerpos (Ac), tambin conocidos como inmunoglobulinas, son un grupo de molculas sricas que producen los linfocitos B. Los diferentes tipos de Ac tienen una estructura bsica comn a todos ellos, pero el sitio por el que se unen al Ag es especfico de cada uno; la parte de la molcula que se une al Ag se denomina regin Fab (fragment antigen binding) mientras que la zona que interacta con otros elementos del SI se denomina regin Fc (algunas clulas del SI tienen sobre su superficie receptores de Fc por lo que si un Ac se une a un patgeno esas clulas tambin pueden unirse a l). La zona de la molcula del Ag a la que se une el Ac se denomina eptopo y una molcula de Ag puede tener varios de ellos por lo que los Ac en realidad son especficos de un eptopo y no de la molcula completa de Ag. Los linfocitos B estn programados para codificar un receptor de superficie especifico de un determinado Ag tras lo cual se multiplican y se diferencian en clulas plasmticas que producen los Ac. Como veremos posteriormente tambin los linfocitos T pueden reconocer Ag aunque no producen Ac. Los linfocitos B y T estn programados genticamente para ser capaces de reconocer especficamente a un determinado Ag antes incluso de haber entrado en contacto con l. Cuando se produce el contacto entre el linfocito y el Ag, los linfocitos que son capaces de reconocerlo empiezan un proceso de proliferacin que conduce en pocos das a la existencia de un nmero suficiente para ocasionar una respuesta inmunitaria que permita la eliminacin del Ag. Es proceso por el que los linfocitos que son capaces de reconocer a un determinado antgeno proliferan se llama seleccin clonal. Una vez producido el contacto inicial con un antgeno determinado, los sucesivos contactos con el mismo antgeno se van a caracterizar por obtener una respuesta mucho ms rpida y enrgica que la inicial debido a que sta da lugar a la produccin de linfocitos de memoria que persisten. El sistema inmunitario dispone adems de diferentes mecanismos de defensa que se denominan genricamente sistemas efectores; ejemplo de ellos son la neutralizacin, la fagocitosis, reacciones citotxicas o la apoptosis celular (muerte celular programada). El sistema linfoide
Las clulas que participan en las respuestas inmunitarias se organizan para formar tejidos y rganos; el conjunto de ellos se denomina sistema linfoide. Existen dos grandes grupos de rganos linfoides, los primarios o centrales y los secundarios o perifricos. En los rganos linfoides primarios se desarrollan y se diferencian los linfocitos dando lugar a clulas maduras a partir de sus precursores (proceso denominado linfopoyesis). En los humanos, la poblacin de linfocitos T madura en el timo y la de linfocitos B en la mdula sea y en el hgado fetal. En estos rganos se adquiere el repertorio de receptores especficos de Ags de tal forma que se presenta tolerancia a los autoantgenos (molculas propias capaces de inducir una respuesta inmune) y cuando viajan a la periferia solo se reconocen Ags extraos. En los rganos linfoides secundarios es necesaria la presencia de macrfagos, clulas presentadoras de antgenos y linfocitos T y B maduros para que se produzca la respuesta inmunitaria. Estos rganos son el bazo, los ganglios linfticos y otros tejidos asociados a la inmunidad de las mucosas, como las amgdalas y las placas de Peyer intestinales; la mdula sea tambin acta como rgano secundario. Las clulas del sistema inmunitario Todas las clulas del SI tienen su origen en clulas madres pluripotenciales de la mdula sea que originan fundamentalmente dos tipos de diferenciacin, la linfoide, que da lugar a los linfocitos, y la mielode, que da origen a los fagocitos. Existen por lo tanto en el SI dos grandes tipos de clulas que intervienen en los procesos de inmunidad. Los fagocitos son capaces de ingerir y degradar antgenos y microorganismos. Dentro de ellos encontramos los fagocitos mononucleares y los neutrfilos polimorfonucleares. El representante genuino de los mononucleares lo constituye la estirpe existente en la sangre perifrica o monocitos que son capaces de emigrar a los tejidos transformndose en los macrfagos tisulares (sistema monocito-macrfago). Los neutrfilos polimorfonucleares o PMN son los leucocitos ms abundantes en sangre perifrica. Adems existen otras clulas, como las clulas presentadoras de antgeno (CPA) a las clulas T, mastocitos, clulas endoteliales, etc. que tambin intervienen en las respuestas inmunitarias y que no pertenecen a ninguno de estos grupos. Los linfocitos Los linfocitos son de dos clases principales, B o T, segn donde se desarrollan; en los humanos, las clulas B se diferencian en la mdula sea y en el hgado fetal y las clulas T en el timo. En estos rganos en los que se diferencian los linfocitos, rganos linfoides primarios, las clulas B y T adquieren la capacidad para reconocer Ags por medio de la adquisicin de receptores de superficie especficos. Existe una tercera clase de linfocitos que no expresan receptores de Ags y que se denominan clulas asesinas naturales (NK, natural killer). Se calcula que en el organismo humano existen del orden de 1012 clulas linfoides y que aproximadamente 109 linfocitos se producen diariamente; la mitad de ellos se renuevan en poco ms de un da, sin embargo otros persisten durante aos e incluso algunos, probablemente, de por vida. Los linfocitos producen molculas de diversa naturaleza que se denominan de un modo general mediadores solubles de la inmunidad. Los principales son los anticuerpos y las citoquinas, pero adems producen diferentes substancias sricas, como el complemento, que actan en procesos inflamatorios. Durante la respuesta inmunitaria las citoquinas transmiten seales entre diferentes tipos celulares; entre sus principales tipos se encuentran los interferones (IFN) que evitan la diseminacin de algunas infecciones vricas, las interleucinas (IL) que fundamentalmente inducen la diferenciacin y multiplicacin de algunas clulas, los factores estimulantes de las colonias (CSF) que intervienen en la diferenciacin y multiplicacin de las clulas madre de la mdula sea, los factores de necrosis tumoral (TNF) o el factor transformador del crecimiento (TGF). Los linfocitos B estn programados para codificar un receptor de superficie especfico de un determinado Ag tras lo cual se multiplican y diferencian en clulas plasmticas que producen Ac. Los linfocitos T tienen diversas funciones. Algunos interactan con las clulas B y los fagocitos mononucleares y se denominan clulas T colaboradoras (clulas Th, de helper); otras destruyen clulas infectadas por agentes intracelulares y se denominan clulas T citotxicas (Tc). La mayora (ms del 90%) de las clulas T son clulas Th.
El sistema ms sencillo, pero simple, de diferenciacin de los linfocitos es su comportamiento tintorial. Sin embargo la expresin de una gran cantidad de molculas de superficie diferentes ha permitido, mediante el empleo de anticuerpos monoclonales especficos, reconocer diferentes poblaciones. Uno de los sistemas desarrollado y empleado en la actualidad es el de denominacin de grupo o CD (cluster designation) que permite la agrupacin de Ac monoclonales de caractersticas similares, aunque puede presentar el inconveniente de que un mismo marcador lo puedan tener lneas celulares diversas o la misma lnea en fases funcionales diferentes. El marcador ms utilizado de las lneas celulares T es el denominado receptor de clulas T (TCR) que las permite separar en dos tipos principales: las clulas T TCR-1 y las clulas T TRC-2; entre el 90-95% de las clulas T expresan TCR-2 y el resto TCR-1. Las clulas T TCR-2 se agrupan a su vez en dos subpoblaciones diferentes: Las que poseen el marcador CD4 que actan como colaboradoras e inductoras de la respuesta inmune (Th) y las que poseen el marcador CD8 que tienen una funcin principalmente citotxica (Tc). Segn expresen o no el marcador se suelen referir con los smbolos + o a continuacin del nmero de CD asignado (pe, CD4+ o CD4-). Las clulas T CD4+ reconocen Ags especficos asociados con molculas del complejo principal de histocompatibilidad (CPH) clase II mientras que las clulas T CD8+ reconocen Ags asociados a molculas CPH de clase I. Solo una pequea fraccin de las clulas T TCR-1 circulantes son CD8+. Las poblaciones T TCR-2+CD4+ se subdividen en dos poblaciones funcionalmente diferentes. De un lado las que ejercen una influencia positiva sobre la respuesta de las clulas T y B (colaboracin) que son CD29+ y que en su mayora expresan una variante del antgeno leucocitario comn CD45 llamada CD45RO y que por lo general se las denomina tambin clulas memoria o CD45RO+. De otro lado las que ejercen la influencia supresora/citotxica de las CD8+ y que expresan un molcula CD45 llamada CD45RA y que por lo general se las denomina tambin clulas ingenuas, vrgenes o CD45RA+. Otras divisiones de las clulas T CD4+ las han agrupado en funcin de las citoquinas que producen hablndose de clones T CD4+ TH1 que secretan IL-2 e IFN gamma y los clones T CD4+ TH2 secretan diferentes interleucinas (IL-4, IL-5, IL-10) que estimulan a las clulas B mientras que las TH1 llevan a cabo funciones relacionadas con la citotoxicidad y la reaccin inflamatoria local. De los expuesto puede deducirse que dentro de la subpoblacin de linfocitos T CD4+ existe una heterogeneidad al menos conceptualmente confusa. La existencia de clulas memoria e ingenuas, que presentaran los marcadores fenotpicos CD45RO+, CD45RA+ o CD29+, no siempre tienen una diferenciacin clara porque existe la transicin clula ingenua a clula memoria. Los clones TH1 y TH2 responden a una concepcin funcional de produccin de ciertas citoquinas por similitud con un sistema murino, pero en humanos no existen marcadores fenotpicos que puedan diferenciar bien estas poblaciones por lo que se emplean diferentes modelos de estudio que no siempre conducen a los mismos datos; adems se piensa que cada subtipo tendra un efector supresor sobre el otro e incluso se ha propuesto la teora de que la inmunodeficiencia del SIDA se debera precisamente a un desequilibrio TH1/TH2 a favor de la poblacin TH2 ya que el VIH infectara fundamentalmente a clones TH1. Las clulas T CD8+, o citotxicas, tambin ese han dividido en diferentes subpoblaciones funcionales que expresan diferentes molculas CD. Tambin se ha descrito una pequea subpoblacin TCR-2 doble negativa (CD4-CD8-). Por otro lado las clulas T TCR-1+ slo constituyen una pequea parte de las clulas T circulantes, abundan ms en las superficies mucosas y epidrmicas, y suelen expresar CD8+ dirigido frente a Ag bacterianos. Se sabe tambin que existen clulas T supresoras (Ts) especficas de Ag aunque no se conoce si constituyen una subpoblacin diferente ya que tanto las clulas T CD4+ y T CD8+ pueden suprimir la respuesta inmunitaria tanto por citotoxicidad directa sobre las CPA como por citocinas supresoras. Los linfocitos B constituyen entre el 5-15% de los linfocitos circulantes y se caracterizan por la presencia de inmunoglobulinas (Ig) de superficie que son sintetizadas por la propia clula y se encuentran en la membrana celular actuando como receptores de Ag. La mayora de los linfocitos B expresan IgM e IgD sobre su superficie y poseen Ag CPH de clase II. La IgM de superficie forma con otras molculas de superficie el llamado complejo receptor de antgenos de las clulas
B (BCR, B cell receptor). Las clulas asesinas naturales constituyen aproximadamente el 15% de los linfocitos circulantes y no expresan receptores de Ag TCR ni BCR. Funcionalmente las clulas NK destruyen clulas tumorales e infectadas por virus. Inmunidad frente a los virus La estructura y complejidad gentica de los virus es variable, desde genomas de ARN con pocos genes hasta complejos genomas ADN con ms de 200 genes, pero en su conjunto, desde un punto de vista simplificador, los virus son un conglomerado de protenas y de cidos nucleicos que necesitan de la maquinaria bioqumica de las clulas del husped que infectan y por lo tanto son parsitos intracelulares estrictos. Existen formas de vida ms simples que los virus, como los viroides que son cido nucleico sin protenas y los priones que son protenas infectivas sin cidos nucleicos. Posiblemente el desarrollo evolutivo del sistema inmunitario de las especies ms evolucionadas ha estado condicionado por su necesidad de adaptarse para producir una respuesta eficaz frente a las infecciones producidas por virus y la respuesta inmune frente a ellas abarca la practica totalidad de los sistemas efectores del SI. Por lo general los virus se unen a las clulas de un husped por medio de receptores especficos que determina el tropismo del virus hacia un determinado tipo de clula o tejido. Una vez dentro de la clula husped se libera el cido nucleico del virus y se produce una transcripcin, sntesis proteica y replicacin del genoma vrico, dando lugar a nuevas partculas vricas (viriones) que se ensamblan y son liberadas infectando nuevas clulas. Segn el tipo de virus este proceso general puede presentar matices diferentes. La capacidad que presentan los virus para infectar a un husped es muy variable y depende tanto del propio microorganismo como del sujeto que sufre el ataque. En la mayora de las infecciones vricas stas se resuelven cuando el husped ha eliminado todos los virus, pero en alguno casos stos pueden quedar en un estado latente y reactivarse bajo determinadas condiciones o bien persistir en su forma infecciosa pese a la respuesta del husped. La primera barrera defensiva frente a las infecciones vricas es la integridad de la superficie corporal; una vez superada sta se ponen en marcha mecanismos defensivos innatos o inespecficos del SI, como las clulas NK, los macrfagos y determinados tipos de interferon (del que se describen tres tipos principales: el IFN-alfa o leucocitario, el IFN-beta o fibroblstico y el IFN-gamma o inmunitario) que estimulan los mecanismos dirigidos a inhibir la replicacin del virus y aumentar la eficacia de la respuesta inmunitaria al inducir un aumento de la expresin de molculas CPH de clases I y II y estimulando las clulas NK. Estas se dirigen al sitio de la infeccin y se activan interviniendo como mediadores de la citotoxicidad celular dependiente de anticuerpos (CMCDA). Al progresar la infeccin se produce la respuesta especfica del SI y aparecen clulas Th, Tc y anticuerpos antivirus que son una barrera fundamental para evitar la diseminacin del virus por la sangre y su transmisin entre clulas. Esta inmunidad mediada por AC presenta mecanismos diversos y puede ser de diferentes tipos; si bien los Ac se pueden generar frente a cualquier protena vrica que se encuentre en la clula infectada los fundamentales son los que se dirigen frente a las glucoprotenas que se expresan en la superficie de las clulas infectadas o de los viriones. Los Ac pueden actuar, solos o con el sistema complemento, frente al virus libre o frente a las clulas infectadas por el virus. Frente al virus libre, el Ac puede bloquear su unin a la clula o bien bloquear la penetracin en la clula o bloquear la eliminacin de la cubierta vrica mientras que con el complemento puede bloquear los receptores del virus. Frente a las clulas infectadas puede ocasionar su lisis, facilitar su fagocitosis o causar CMCDA por parte de las clulas NK o los macrfagos. Las clulas T ejercen diferentes funciones en la respuesta de inmunidad frente a los virus; la mayora de las respuestas de AC son timodependientes por lo que precisan la colaboracin de las clulas T CD4+ que adems activan clulas T citotxicas CD8+ y macrfagos. Entre los mecanismos que utilizan los virus para escapar a la accin del SI, la variacin antignica es el sistema defensivo ms eficaz para evitar ser reconocidos. Por lo general consiste en la mutacin de regiones proteicas contra las que estn dirigidos los AC y en estos casos la inmunidad humoral frente a las enfermedades que provocan solo persiste hasta que son capaces de desarrollan una cepa viral nueva lo que ocasiona que las vacunas que se pueden lograr no son eficaces a largo plazo. Otros de los mecanismos por los
que los virus pueden eludir los mecanismos de defensa del SI son el ocasionar exclusivamente infecciones intracelulares, la produccin de molculas que compiten o inhiben ciertas enzimas, o la formacin de compuestos homlogos a los receptores de las citocinas o a ellas mismas.
LAS ENFERMEDADES INFECCIOSAS Y EL SISTEMA INMUNE. En estos tiempos, al final del siglo XX, la humanidad pareca haber dominado a las enfermedades infecciosas. Desde los hallazgos de Koch, Pasteur y colaboradores hasta la elaboracin e implementacin de los novsimos antibiticos todo pareca ser victoria. Pero la aparicin de la resistencia bacteriana y, posteriormente del VIH, pusieron un signo de interrogacin a esa creencia. Pronto fue aparente que no todo dependa de la tecnologa; en el fondo estaba la accin del sistema inmune.Resulta instructivo describir el conocimiento corriente de las acciones del sistema inmune en relacin a las diversas posibilidades de infeccin: diferentes tipos de agente biolgico y distintos lugares de ubicacin del mismo. Conviene no olvidar que, en un sentido amplio, cada uno de los componentes del sistema parece estar dirigido contra agentes que afectan un nicho particular en el organismo. Por ejemplo los anticuerpos son especialmente efectivos en la destruccin de las bacterias que viven fuera de las clulas humanas, mientras que los linfocitos T CD4 son especialmente importantes en la destruccin de parsitos intracelulares; a pesar de estas especializaciones la respuesta inmune involucra a todos los componentes del sistema que actan armnicamente. Cada componente se involucra, directa o indirectamente, en repeler la agresin y, en este aspecto adquieren nuevos roles, por ejemplo los linfocitos T CD4 han ganado el nombre de "cooperadores" o "auxiliares" porque producen y secretan sustancias que amplifican y controlan virtualmente a todos los aspectos de la inmunidad. LA LUCHA CONTRA LOS PATOGENOS EXTRACELULARES. La neumona es un padecimiento inflamatorio del parnquima pulmonar; en su forma mas comn es causada por la bacteria S. pneumoniae, el neumococo. Esta bacteria esfrica es capaz de entrar hasta la luz alveolar en donde producen una reaccin inflamatoria que daa a los tejidos y dificulta el intercambio gaseoso. Si la infeccin no es tratada oportuna y eficazmente se pueden desarrollar complicaciones y cierto nmero de pacientes morirn. Los neumococos viven en la luz alveolar donde se encuentran al alcance de los fagocitos: neutrfilos, macrfagos y otros;empero los neumococos escapan a la deteccin y a la fagocitosis mediante un curioso artilugio: la creacin de una envoltura externa formada por mltiples molculas de azcares, cpsula polisacrida que evita el acercamiento y reconocimiento de los fagocitos. El artilugio defensivo de los neumococos es eficaz para evitar la fagocitosis y le permite a la bacteria establecerse en los pulmones sin ser destruida, pero la cpsula de polisacridos no es capaz de defender a la bacteria del ataque de los anticuerpos producidos por los linfocitos B. Los linfocitos B presentan en la superficie de sus membranas receptores para todos los antgenos (sustancias extraas) concebibles, pero cada clula en particular nicamente posee receptores para uno solo de esos antgenos, como existen miles de millones de stas clulas es casi seguro que alguno de estos receptores pueda embonarse, como una llave con la cerradura, a las molculas de la cpsula bacteriana; por la peculiar estructura de esta cpsula bacteriana, los mismos antgenos aparecen regularmente y pueden adherirse con muchos receptores de la clula B. Este factor resulta decisivo para la activacin del linfocito B. Los linfocitos B activados, por el proceso descrito y las citoquinas provenientes de los linfocitos T auxiliares, desencadenan una actividad frentica que las lleva, en primer lugar a dividirse muchas veces multiplicando, de esta manera el nmero de receptores que pueden ligarse al antgeno bacteriano. En un momento posterior, estos receptores se liberarn de la superficie del linfocito B y, desde este momento, podemos llamarlos "anticuerpos". Los anticuerpos se fijan a los antgenos de la cpsula polisacrida bacteriana y mejoran la habilidad de los macrfagos para fagocitarla con mecanismos como el siguiente: los anticuerpos formados por Inmunoglobulina G (IgG) presentan una regin denominada Fc-gamma; en los macrfagos y otros fagocitos existen receptores para esta regin y, cuando la misma es reconocida el macrfago recibe la seal para fagocitar la partcula. Puede decirse que, los anticuerpos prestan a los macrfagos el apoyo necesario para
consumar la fagocitosis. Por otro lado los anticuerpos unidos a la cpsula bacteriana la hacen mas vulnerable a la actividad de una cascada de enzimas circulantes a las que conocemos como "sistema del complemento" que no solamente ataca a la bacteria sino que tambin provee de una seal de retroalimentacin para que las clulas B produzcan mas anticuerpos. El mecanismo que acabamos de describir en muchos aspectos es el mas simple en el repertorio del sistema inmune y bsicamente consiste en una carrera de produccin entre las bacterias y los anticuerpos. La resistencia la infeccin que desarrollan las personas vacunadas depende bsicamente de la produccin de anticuerpos. Los anticuerpos capaces de neutralizar las infecciones virales se conocen como anticuerpos neutralizantes (AcN) y producen diversos efectos: como la inhibicin de los procesos de fijacin, penetracin o desnudamiento del virus, aglomeran a los viriones o bien provocan su opsonizacin. Como los virus son parsitos intracelulares es poco el tiempo en que se encuentran al alcance de los anticuerpos y, en consecuencia, estos tiene un efecto muy pequeo en la defensa del organismo
LAS INFECCIONES INTRACELULARES. Muchos parsitos: protozoarios como la Leishmania, bacterias como las micobacterias y todos los virus son capaces de establecerse, multiplicarse y prosperar en el interior de nuestras clulas; Leishmania y las micobacterias se establecen dentro de vacuolas o el retculo endoplsmico rugoso, mientras que los virus lo hacen en el citosol. Los mecanismos de ataque del sistema inmunolgico son distintos en cada caso. En los casos de ubicacin intracelular el parsito invasor se expone solo durante breves momentos al ataque de los anticuerpos y el complemento, mientras pasan de una clula parasitada a la otra. Tomemos el caso de las micobacterias; estas bacterias producen la tuberculosis y la lepra, dos enfermedades realmente temibles. M. tuberculossis carece de motilidad propia y, en cuanto entran los bacilos de este gnero entran al organismo son fagocitados por los macrfagos y, dentro de estas clulas, son colocados en vacuolas para ser expuestas a un bao enzimtico que, en condiciones naturales, puede destruir a la mayor parte de las bacterias. Sin embargo estos bacilos no son afectados y prosperan dentro de este singular nicho ecolgico; pueden replicarse y parasitar al fagocito hasta que agotan sus reservas y la clula hospedera muere. La lucha contra estos parsitos depende de las molculas del Complejo Principal de Histocompatibilidad (CPH), un grupo de molculas producidas en el interior de la clula ligadas a antgenos del parsito; regularmente estas molculas migran hacia la membrana donde los antgenos son expuestos al reconocimiento de los linfocitos T auxiliares (CD4) que, de esta manera, se percatan de la presencia de una infeccin intracelular. Como en el caso de las clulas B, tambin estos linfocitos poseen la diversidad suficiente para "reconocer" cualquier combinacin entre las molculas del CPH y un antgeno del parsito. En la superficie de estas clulas parasitadas puede encontrarse otra molcula proteica, la B7. Esta proteina B7 es reconocida por un receptor complementario en la superficie de los linfocitos CD4 que se activan al recibir esta doble seal, o se anergizan en el caso de no recibirla. Los linfocitos T cooperadores activados asumen la tarea de montar la respuesta inmunolgica y realizan este trabajo liberando numerosas sustancias, las citoquinas, que regulan la accin de las otras clulas integrantes del sistema inmune. Una de estas sustancias, el interfern gamma incita al macrfago a producir otras citoquinas, como el factor de necrosis tumoral y diversas sustancias como el xido ntrico y radicales txicos del oxgeno que pueden destruir a las bacterias. Sin embargo esta no es la nica posibilidad de respuesta de los linfocitos T auxiliares puesto que, en lugar de producir interfern gamma sino que, en respuesta al estmulo producen otras citoquinas: la interleuquina-4 y la interleuquina-10 que bloquean la respuesta inducida por elinterfern gamma!. La eleccin del tipo de citoquinas que producirn las clulas T resulta el incidente crtico para conocer el curso de la enfermedad. Desde este punto de vista se ha intentado aclarar la patogenia de la lepra; como es sabido existen dos formas de la enfermedad: la lepromatosa y la tuberculoide. En la lepra tuberculoide los linfocitos T producen interfern gamma y se monta una vigorosa respuesta inmune; por el contrario, en la lepra lepromatosa la respuesta inmune es dbil, dominada por la produccin de interleuquina-4 y las lesiones presentan vastas lesiones conteniendo grandes poblaciones de bacilos intercelulares. Los hechos aqu descritos han llevado a la suposicin de que es posible aumentar la eficacia de la respuesta
inmune mediante la administracin de anticuerpos monoclonales dirigidos contra la interleuquina-4.
LA REPUESTA A LOS VIRUS. A pesar de que tambin se encuentran en el interior de las clulas los virus plantean problemas diferentes al sistema inmune. Los virus se encuentran en el citosol y llegan a fusionarse tan ntimamente con la clula que podran parecer partes de la misma. Sin embargo el CPH puede arreglrselas para arrancar antgenos virales y llevarlos dentro de vesculas secretorias a la membrana celular, donde examinados y reconocidos por linfocitos T, pero en este caso CD8. Las clulas citolticas (NK=natural killers= asesinos naturales) son grandes linfocitos granulosos capaces de fijarse a las clulas diana infectadas y secretar, a continuacin, molculas citotxicas dentro de vesculas granulosas azurfilas. Esta variedad de linfocitos producen perforina y otras enzimas que destruyen la membrana celular de las clulas infectadas o son capaces de activar determinados mecanismos de autodestruccin celulares produciendo una especie de suicidio celular conocido como apoptosis. Los linfocitos T CD8 se han convertido en linfocitos asesinos capaces de liberar interfern gamma y factor necrosante de los tumores. Estas molculas limitan la replicacin viral puesto que destruyen las clulas antes de que las partculas virales completen su maduracin y constituyen la defensa mas precoz frente a las infecciones virales pues su mxima accin ocurre a los 2-3 das despus de la iniciacin de la infeccin. La funcin de estos NK se potencia con los interferones y se han demostrado en casos de sarampin y Virus de Epstein-Barr. LA GUERRA DE TERRENO ARRASADO. Los mecanismos del sistema de inmunidad mediados por los linfocitos asesinos puede ser tan devastadora que dae al hospedero y, en estos casos, el remedio puede ser peor que la enfermedad puesto que muchos virus no son "citopticos", es decir, no daan a la clula, pero la respuesta inmune puede hacerlo. Esto es evidente en enfermedades como la hepatitis viral B; el virus no causa dao en los hepatocitos, pero la respuesta inmune destruye un enorme nmero de estas tiles clulas. El organismo parece controlar estos casos cuando existe un nmero abrumador de invasores no citopticos; las clulas CD8 sufren una gran activacin, una actividad exhuberante y luego mueren dejando una brecha inmunolgica. La inmunopatologa producida por los virus puede resumirse de la manera siguiente: 1. Formacin de anticuerpos policlonales contra antgenos no relacionados con el virus originador. 2. Formacin de anticuerpos por reaccin cruzada contra estructuras normales del hospedero con estructuras moleculares similares. 3. Formacin de inmunocomplejos que se precipitan en la membrana basal de algunos epitelios desde donde atraen a los mediadores de inflamacin y producen destruccin tisular. 4. Formacin de autoanticuerpos contra linfocitos, plaquetas, mielina, etc. Los ejemplos sealados hasta aqu ilustran esquemticamente la accin del sistema inmune, elegante pero imperfecto; que la mayora de las veces nos protege de los agresores externos, pero otras veces puede llegar a favorecerlos. La aclaracin de sus mltiples mecanismos de accin nos capacitarn para afinar su eficacia y aprovechar sus notables propiedades.
REACCIONES DE HIPERSENSIBILIDAD
Los mecanismos de inmunidad normalmente nos protegen aislando al yo de cualesquiera otros fenmenos del "no yo"; sin embargo pueden fracasar de tres maneras distintas: 1) Los mecanismos de inmunidad fracasan por incapacidad de frenar a los invasores. A este grupo pertenecen la mayora de los padecimientos por inmunodeficiencia, congnitas y adquiridos. 2) Los mecanismos de defensa se vuelven incapaces de distinguir entre el yo y el no yo y atacan a los tejidos corporales y provocan enfermedades por autoinmunidad como la Diabetes mellitus tipo I, etc. 3) Los mecanismos de defensa reaccionan con exageracin y producen una respuesta que, mas que defender, ataca a las estructuras del hospedero. A stos fenmenos se les llama Reacciones de hipersensibilidad y se han clasificado (Gell y Combs) en cuatro tipos:
Tipo I: reacciones anafilcticas: Este tipo de reacciones se produce cuando un antgeno estimula la produccin de una especie particular de anticuerpo: el anticuerpo reagnico (en los seres humanos formado por Ig. E); este anticuerpo es capaz de fijarse a las clulas cebadas (mastocitos) y a los basfilos (cierto tipo de leucocitos) producindose un proceso que se llama sensibilizacin. Tiempo despus, una reaccin entre el antgeno (libre) y el anticuerpo ( fijado a las clulas cebadas), produce la desgranulacin de estas clulas que contienen poderosos mediadores como la histamina, la serotonina, la sustancia de reaccin lenta de la anafilaxis y la heparina. La liberacin de estas sustancias ocurre con rapidez y por esta razn a este fenmeno tambin se le llama una reaccin de hipersensibilidad inmediata. Los sndromes clnicos abarcan la anafilaxia generalizada, la fiebre del heno, la rinitis alrgica y el asma extrnseca. En su produccin interviene la IgE y la desgranulacin mastocitaria. Las pruebas cutneas dan una reaccin inmediata de ppula y eritema, producidas por un cambio rpido de la permeabilidad microvascular en respuesta a la liberacin local de mediadores vasoactivos como la histamina. No intervienen otrs inmunoglubulinas, ni el complemento ni otras clulas inmunoefectoras. Se habla de un choque anafilctico para describir a las reacciones anafilcticas graves que ponen en peligro la vida; estas reacciones se parecen mucho a las que producira una dosis de histamina inyectada por va endovenosa: hipotensin grave por intensa vasodilatacin perifrica y constriccin bronquiolar. Este fenmeno se desencadena con mucha rapidez, prcticamente de inmediato en cuanto penetra un antgeno al torrente sanguneo; en los seres humanos este peligro surge al intentar alguna medida teraputica como administrar un suero, aplicar penicilina, medios de contraste iodados, etc.. No siempre se produce choque anafilctico cuando ocurre una exposicin al antgeno; cuando ste antgeno se pone en contacto con la circulacin de tal manera que aumente rpidamente su concentracin sangunea. En estos casos se producen reacciones anafilcticas locales como: *La fiebre del heno, inflamacin anafilctica tpica de las vas respiratorias superiores despus de inhalar un antgeno. *El asma bronquial alrgica que afecta al msculo liso de los bronquiolos y a las secreciones. Figura 1: Mastocito y mastocisto degranulado *Vmito y diarrea al ingerir alimentos con antgenos a los que se est sensibilizado. *Urticaria en la piel, al entrar en contacto con un antgeno. A este tipo de reacciones locales por hipersensibilidad suele llamrseles alergia y, a las sustancias que desencadenan las reacciones alrgicas se les llama alergenos. Entre los alergenos mas comunes podemos sealar: 1) En la casa: plenes, mohos, partes de insectos, caros, plumas y pieles. 2) En los alimentos: leche, huevo, pescado y moluscos. 3) En el campo: picadura de insectos.
4) En la medicina: algunos frmacos como la penicilina, sulfamdicos, salicilatos, medios de contraste iodados, etc. Las reacciones alrgicas son una caracterstica de las infestaciones por helmintos. La reaccin alrgica mas espectacular es la que sigue a la ruptura de un quiste hidatdico ya que la lenta filtracin de los antgenos del gusano asegura que los mastocitos del enfermo estarn sensibilizados con una IgE especfica y la masiva llegada de antgeno tras la ruptura del quiste puede producir una reaccin mortal por anafilaxis.. Tambin se pueden presentar estas reacciones en las infestaciones por A lumbricoides La susceptibilidad individual para los antgenos es altamente individual y se desconoce el por qu algunas personas la desarrollan mientras que otras personas nunca la presentan. Se llama atopa a la alta susceptibilidad de una persona para sufrir este tipo de enfermedades; la base de la atopa es gentica con genes determinantes de penetrancia incompleta. Cuando una persona presenta alguna alergia aumentan sus posibilidades de sufrir otra reaccin de este tipo. Las reacciones de este tipo pueden ser bloqueadas por grandes cantidades de antihistamnicos (que bloquean a los receptores) o bien por corticosteroides que inhiben la liberacin de las enzimas que intervienen en la formacin de mediadores.
Tipo II. Reacciones citolticas o citotxicas. En este tipo de reaccin de hipersensibilidad se produce la reaccin de un anticuerpo circulante de clase IgG, IgM o IgA con un antgeno asociado a la membrana celular. El ejemplo mas comprensible en este tipo de reacciones se presenta en los eritrocitos cuando se administra una transfusin de sangre incompatible; en este caso se forma un complejo antgeno-anticuerpo, fijacin del complemento y lisis de la clula. En el sistema respiratorio ocurre una de estas reacciones en el llamado "Sndrome de Goodpasture" en el cual el excremento de palomas puede desencadenar la destruccin de los epitelios del rin (manifestado por hematuria) y pulmn (manifestado por hemoptisis). Tipo III. Reacciones de complejos (precipitina txica). Aqu el dao lo produce la unin de un antgeno soluble (por ejemplo toxinas bacterianas) con anticuerpos circulantes; normalmente estos "complejos" son llevados al bazo donde desaparecen de la circulacin; pero en condiciones como el exceso de antgeno se forma un tipo de complejos que se precipita en algunos sitios como el revestimiento endotelial de los vasos sanguneos o las membranas glomerulares del rin. Ciertos tipos de Asma Bronquial obedecen a este mecanismo , lo mismo que la enfermedad de Arthus y la enfermedad del suero. Estas reacciones se demoran mas en aparecer que lo que hacen las reacciones de tipo I y no responden a los corticoides. Tipo IV. Reacciones de hipersensibilidad retardada (mediada por clulas). Estas reacciones son producidas por la presencia de linfocitos T "asesinos" que intentan destruir al antgeno; caractersticamente se presentan entre 12 y 24 horas despus de la inyeccin intracutnea de un antgeno (como ocurre en la prueba de la tuberculina o PPD). La reaccin de las clulas T "asesinas" con el antgeno produce la muerte de dichas clulas y este es el factor que desencadena una respuesta inflamatoria. La patogenia de la tuberculosis pulmonar obedece a este mecanismo de hipersensibilidad, lo mismo ocurre en la lepra y otras enfermedades micticas y parasitarias as como en las dermatitis por contacto. Desde el punto de vista de los mdicos, el aspecto mas importante de estas reacciones de tipo IV consiste en la formacin de un granuloma. La accin de las citocinas determinar la evolucin de ese granuloma (que es diferente en la tuberculosis y en la lepra).
Anda mungkin juga menyukai
- Calendario de Estudio ENARMDokumen14 halamanCalendario de Estudio ENARMElsa Moreno83% (6)
- Fundamentos de La Ventilacion MecanicaDokumen62 halamanFundamentos de La Ventilacion MecanicaAdriana OlivaresBelum ada peringkat
- Sistema Endocrino y Reproductor - Cursos Crash PDFDokumen288 halamanSistema Endocrino y Reproductor - Cursos Crash PDFUleonel86% (7)
- Fisiología de La Coagulación Modificada Junio 2013Dokumen46 halamanFisiología de La Coagulación Modificada Junio 2013alyzon mello montejoBelum ada peringkat
- IslasVargas - Leticia - Aparato Digestivo - RepositorioDokumen10 halamanIslasVargas - Leticia - Aparato Digestivo - RepositorioAdriana OlivaresBelum ada peringkat
- 10 Ictericia y ColestasisDokumen12 halaman10 Ictericia y ColestasisYanita Avila ClazngBelum ada peringkat
- Proyecto Material de Curacion 2016Dokumen201 halamanProyecto Material de Curacion 2016Adriana OlivaresBelum ada peringkat
- ColeeeeeeeeDokumen3 halamanColeeeeeeeeAdriana OlivaresBelum ada peringkat
- 16 Tecnicas Imagen 0 PDFDokumen27 halaman16 Tecnicas Imagen 0 PDFGil MartinezBelum ada peringkat
- Cuaderno de NudosDokumen26 halamanCuaderno de NudosAdriana OlivaresBelum ada peringkat
- Fisiología de La Coagulación Modificada Junio 2013Dokumen46 halamanFisiología de La Coagulación Modificada Junio 2013alyzon mello montejoBelum ada peringkat
- Triptico Grupo Platino 5Dokumen73 halamanTriptico Grupo Platino 5Adriana OlivaresBelum ada peringkat
- Calendario Platino 5Dokumen8 halamanCalendario Platino 5Adriana OlivaresBelum ada peringkat
- 30 Mecanismos Locales de ProtecciónDokumen6 halaman30 Mecanismos Locales de ProtecciónAdriana OlivaresBelum ada peringkat
- Caso Clinico 3Dokumen17 halamanCaso Clinico 3Adriana OlivaresBelum ada peringkat
- BRONQUIOLITISDokumen3 halamanBRONQUIOLITISAdriana OlivaresBelum ada peringkat
- Traduccion Del Video Anticurpos MediadoresDokumen1 halamanTraduccion Del Video Anticurpos MediadoresAdriana OlivaresBelum ada peringkat
- ESCABIASISDokumen46 halamanESCABIASISAdriana OlivaresBelum ada peringkat
- Cap 2 Act 6Dokumen3 halamanCap 2 Act 6Adriana OlivaresBelum ada peringkat
- Cuadro de UroDokumen1 halamanCuadro de UroAdriana OlivaresBelum ada peringkat
- Boca Parte 1Dokumen19 halamanBoca Parte 1Adriana OlivaresBelum ada peringkat
- Transporte de Hemoglobina y Equilibrio Ácido BaseDokumen1 halamanTransporte de Hemoglobina y Equilibrio Ácido BaseAdriana OlivaresBelum ada peringkat
- DIFUSIÓNDokumen1 halamanDIFUSIÓNAdriana Olivares100% (1)
- Exam Diges 1Dokumen4 halamanExam Diges 1Adriana OlivaresBelum ada peringkat
- NeumoniaDokumen23 halamanNeumoniaAdriana OlivaresBelum ada peringkat
- 29 OlfatoDokumen3 halaman29 OlfatoAdriana OlivaresBelum ada peringkat
- 28 FonaciónDokumen5 halaman28 FonaciónAdriana OlivaresBelum ada peringkat
- 22 ControlDokumen3 halaman22 ControlAdriana OlivaresBelum ada peringkat
- Semana 13 Exploración Osteomioarticular IiiDokumen59 halamanSemana 13 Exploración Osteomioarticular IiiAdriana OlivaresBelum ada peringkat
- Sanación Pránica Violeta-1Dokumen21 halamanSanación Pránica Violeta-1cmuñoz_568442100% (1)
- FIEBREDokumen11 halamanFIEBREJuan Pablo SempéBelum ada peringkat
- Antecedentes Personales PatológicosDokumen1 halamanAntecedentes Personales PatológicosiQueTetoBelum ada peringkat
- Resumen II Bloque - MicroDokumen182 halamanResumen II Bloque - Microgabriela loarcaBelum ada peringkat
- ANTIHISTAMINICOSDokumen29 halamanANTIHISTAMINICOSBeatriz MarquezBelum ada peringkat
- Farmacologia G3Dokumen33 halamanFarmacologia G3KEHILY PAULINA PINARGOTE LEONBelum ada peringkat
- Dieta ReumaDokumen3 halamanDieta ReumaSonia Vidal SanzanaBelum ada peringkat
- Manual SemiologiaDokumen24 halamanManual SemiologiaNicolásBelum ada peringkat
- Cabinas de Bioseguridad - ABDokumen86 halamanCabinas de Bioseguridad - ABJarod Varela100% (2)
- OftalmologiaDokumen48 halamanOftalmologiaDnns PrietBelum ada peringkat
- RFRRDDokumen12 halamanRFRRDAnGie AmesquitaBelum ada peringkat
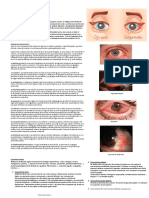
- Conjuntivitis - ResumenDokumen2 halamanConjuntivitis - ResumenLuccia CaracoBelum ada peringkat
- Pintura Jotamastic 90 Comp ADokumen14 halamanPintura Jotamastic 90 Comp AgermanBelum ada peringkat
- Fitoterapia y La Cosmética NaturalDokumen5 halamanFitoterapia y La Cosmética Naturalalejahebe0829Belum ada peringkat
- Proyecto Colonia de BebeDokumen24 halamanProyecto Colonia de BebeJoseph Avila Alvarez0% (2)
- MP - Respuesta Inmune. 2022Dokumen13 halamanMP - Respuesta Inmune. 2022CÉSAR EMMANUEL LANDEROS CORONABelum ada peringkat
- Hempadur 8567511150 Es-EsDokumen11 halamanHempadur 8567511150 Es-EsJairo ArroyoBelum ada peringkat
- Colitis Alérgica Eosinofílica.Dokumen9 halamanColitis Alérgica Eosinofílica.entredosmundosBelum ada peringkat
- Ficha de AlumnoDokumen36 halamanFicha de AlumnoVc CamachoBelum ada peringkat
- Copia de Formato Ficha TecnicaDokumen3 halamanCopia de Formato Ficha TecnicaMauricio Orlando Vanegas CasadiegoBelum ada peringkat
- AblactaciónDokumen9 halamanAblactaciónHeriberto Juarez100% (1)
- Consentimiento Informado 2018 PDFDokumen3 halamanConsentimiento Informado 2018 PDFMaria Esther Torres DiazBelum ada peringkat
- Ejemplos Metodo CientificoDokumen6 halamanEjemplos Metodo CientificoJorge Raul Puello SilvaBelum ada peringkat
- Cuidado Integral Del Paciente PediatricoDokumen24 halamanCuidado Integral Del Paciente Pediatricokatherine milagros rodriguez montañezBelum ada peringkat
- Conjuntivitis BacterianaDokumen9 halamanConjuntivitis BacterianaKristal RamirezBelum ada peringkat
- InmunotoxicidadDokumen8 halamanInmunotoxicidadAlberto Junior Quispe YayaBelum ada peringkat
- Inmunidad AdquiridaDokumen8 halamanInmunidad AdquiridaJefferson Loor0% (1)
- AlérgenosDokumen2 halamanAlérgenosMiguel GallegosBelum ada peringkat
- PE Dermovate CremaDokumen14 halamanPE Dermovate CremaMaria CanalesBelum ada peringkat
- Evidencia 1 Matriz Determinar Las Eventualidades A Presentarse Durante Recorrido Grupo 5Dokumen15 halamanEvidencia 1 Matriz Determinar Las Eventualidades A Presentarse Durante Recorrido Grupo 5Lucy Liliana Vargas Bermúdez100% (1)