Anda mungkin juga menyukai
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- The Telepresence Add-In The Trackit! Add-InDokumen60 halamanThe Telepresence Add-In The Trackit! Add-InemadhsobhyBelum ada peringkat
- شبكة الرعاية الصحية.xlsx - NW - EnglishDokumen111 halamanشبكة الرعاية الصحية.xlsx - NW - EnglishemadhsobhyBelum ada peringkat
- 5 Homes Around The World Oxford Read and Disc PDFDokumen30 halaman5 Homes Around The World Oxford Read and Disc PDFemadhsobhy100% (1)
- Stage Management Using The Stage ManagerDokumen23 halamanStage Management Using The Stage ManageremadhsobhyBelum ada peringkat
- SliceViewer and VoxelViewer - enDokumen55 halamanSliceViewer and VoxelViewer - enemadhsobhyBelum ada peringkat
- Weld Measurement enDokumen19 halamanWeld Measurement enemadhsobhyBelum ada peringkat
- Analysis: The Telepresence Add-In The Telepresence Add-InDokumen32 halamanAnalysis: The Telepresence Add-In The Telepresence Add-InemadhsobhyBelum ada peringkat
- Analysis: Add - in Intercept CountingDokumen31 halamanAnalysis: Add - in Intercept CountingemadhsobhyBelum ada peringkat
- Virtual Stage enDokumen12 halamanVirtual Stage enemadhsobhyBelum ada peringkat
- User's Manual Inclusion InspectorDokumen74 halamanUser's Manual Inclusion InspectoremadhsobhyBelum ada peringkat
- Image Sequence Processing - enDokumen27 halamanImage Sequence Processing - enemadhsobhyBelum ada peringkat
- Particle Inspector enDokumen95 halamanParticle Inspector enemadhsobhyBelum ada peringkat
- IX enDokumen42 halamanIX enemadhsobhyBelum ada peringkat
- User's Guide FilterinspectorDokumen57 halamanUser's Guide FilterinspectoremadhsobhyBelum ada peringkat
- Cast Iron: ManualDokumen24 halamanCast Iron: ManualemadhsobhyBelum ada peringkat
- Diffraction Image Analysis 2Dokumen16 halamanDiffraction Image Analysis 2emadhsobhyBelum ada peringkat
- Coating enDokumen12 halamanCoating enemadhsobhyBelum ada peringkat
- EFTEM enDokumen158 halamanEFTEM enemadhsobhyBelum ada peringkat
- BX enDokumen46 halamanBX enemadhsobhyBelum ada peringkat
- Step by Step ITEM - enDokumen233 halamanStep by Step ITEM - enemadhsobhyBelum ada peringkat
- ASAC enDokumen33 halamanASAC enemadhsobhyBelum ada peringkat
- Chain Measurement enDokumen33 halamanChain Measurement enemadhsobhyBelum ada peringkat
- MegaViewIII Installation Guide - enDokumen56 halamanMegaViewIII Installation Guide - enemadhsobhyBelum ada peringkat
- Automatic Caliper Tool - enDokumen47 halamanAutomatic Caliper Tool - enemadhsobhyBelum ada peringkat
- Analysis: User'S Guide 3D-ProcessingDokumen27 halamanAnalysis: User'S Guide 3D-ProcessingemadhsobhyBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Chapter13 PDFDokumen34 halamanChapter13 PDFAnastasia BulavinovBelum ada peringkat
- Proceeding of Rasce 2015Dokumen245 halamanProceeding of Rasce 2015Alex ChristopherBelum ada peringkat
- Outdoor Air Pollution: Sources, Health Effects and SolutionsDokumen20 halamanOutdoor Air Pollution: Sources, Health Effects and SolutionsCamelia RadulescuBelum ada peringkat
- DarcDokumen9 halamanDarcJunior BermudezBelum ada peringkat
- Sba 2Dokumen29 halamanSba 2api-377332228Belum ada peringkat
- Plaza 66 Tower 2 Structural Design ChallengesDokumen13 halamanPlaza 66 Tower 2 Structural Design ChallengessrvshBelum ada peringkat
- A Case Study of Coustomer Satisfaction in Demat Account At: A Summer Training ReportDokumen110 halamanA Case Study of Coustomer Satisfaction in Demat Account At: A Summer Training ReportDeepak SinghalBelum ada peringkat
- Ito Na Talaga Yung FinalDokumen22 halamanIto Na Talaga Yung FinalJonas Gian Miguel MadarangBelum ada peringkat
- Project Scheduling and TrackingDokumen47 halamanProject Scheduling and TrackingArun VinodhBelum ada peringkat
- Duavent Drug Study - CunadoDokumen3 halamanDuavent Drug Study - CunadoLexa Moreene Cu�adoBelum ada peringkat
- MASONRYDokumen8 halamanMASONRYJowelyn MaderalBelum ada peringkat
- Sindi and Wahab in 18th CenturyDokumen9 halamanSindi and Wahab in 18th CenturyMujahid Asaadullah AbdullahBelum ada peringkat
- Leak Detection ReportDokumen29 halamanLeak Detection ReportAnnMarie KathleenBelum ada peringkat
- 8.ZXSDR B8200 (L200) Principle and Hardware Structure Training Manual-45Dokumen45 halaman8.ZXSDR B8200 (L200) Principle and Hardware Structure Training Manual-45mehdi_mehdiBelum ada peringkat
- Laboratory Manual (CIV 210) Engineering Surveying (2018-19) (For Private Circulation Only)Dokumen76 halamanLaboratory Manual (CIV 210) Engineering Surveying (2018-19) (For Private Circulation Only)gyanendraBelum ada peringkat
- Service Manual: SV01-NHX40AX03-01E NHX4000 MSX-853 Axis Adjustment Procedure of Z-Axis Zero Return PositionDokumen5 halamanService Manual: SV01-NHX40AX03-01E NHX4000 MSX-853 Axis Adjustment Procedure of Z-Axis Zero Return Positionmahdi elmay100% (3)
- CH 1 - Democracy and American PoliticsDokumen9 halamanCH 1 - Democracy and American PoliticsAndrew Philip ClarkBelum ada peringkat
- Organization and Management Module 3: Quarter 1 - Week 3Dokumen15 halamanOrganization and Management Module 3: Quarter 1 - Week 3juvelyn luegoBelum ada peringkat
- Arens - Auditing and Assurance Services 15e-2Dokumen17 halamanArens - Auditing and Assurance Services 15e-2Magdaline ChuaBelum ada peringkat
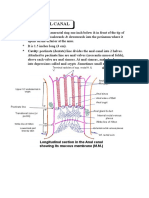
- Anatomy Anal CanalDokumen14 halamanAnatomy Anal CanalBela Ronaldoe100% (1)
- Corrosion Fatigue Phenomena Learned From Failure AnalysisDokumen10 halamanCorrosion Fatigue Phenomena Learned From Failure AnalysisDavid Jose Velandia MunozBelum ada peringkat
- Rule 113 114Dokumen7 halamanRule 113 114Shaila GonzalesBelum ada peringkat
- Lesson 5 Designing and Developing Social AdvocacyDokumen27 halamanLesson 5 Designing and Developing Social Advocacydaniel loberizBelum ada peringkat
- Work ProblemsDokumen19 halamanWork ProblemsOfelia DavidBelum ada peringkat
- Business Analytics Emphasis Course GuideDokumen3 halamanBusiness Analytics Emphasis Course Guidea30000496Belum ada peringkat
- Harper Independent Distributor Tri FoldDokumen2 halamanHarper Independent Distributor Tri FoldYipper ShnipperBelum ada peringkat
- Lesson 6 ComprogDokumen25 halamanLesson 6 ComprogmarkvillaplazaBelum ada peringkat
- Costbenefit Analysis 2015Dokumen459 halamanCostbenefit Analysis 2015TRÂM NGUYỄN THỊ BÍCHBelum ada peringkat
- Classifications of AssessmentsDokumen11 halamanClassifications of AssessmentsClaire CatapangBelum ada peringkat
- Movie Piracy in Ethiopian CinemaDokumen22 halamanMovie Piracy in Ethiopian CinemaBehailu Shiferaw MihireteBelum ada peringkat