Anda mungkin juga menyukai
- A Replicação Do DNA É Semi-ConservativaDokumen3 halamanA Replicação Do DNA É Semi-ConservativaEduarda QueirozBelum ada peringkat
- Genética Médica para Médicos e Estudantes de Medicina: OncologiaDari EverandGenética Médica para Médicos e Estudantes de Medicina: OncologiaBelum ada peringkat
- Questões Subjetivas Parte C e D Do TEXTO 2 A REPLICAÇÃO DO DNADokumen5 halamanQuestões Subjetivas Parte C e D Do TEXTO 2 A REPLICAÇÃO DO DNAcadsus susBelum ada peringkat
- Apostila - 4 - Replicação Do DNA - e - Transcrição-Do DNA em RNA - (Atualizada)Dokumen15 halamanApostila - 4 - Replicação Do DNA - e - Transcrição-Do DNA em RNA - (Atualizada)GEORGIA ANTONIA FERNANDES DOMINICINIBelum ada peringkat
- Professorferretto Professorferretto: Questão B0260: CDokumen1 halamanProfessorferretto Professorferretto: Questão B0260: CGabriel CordeiroBelum ada peringkat
- GENÉTICA HUMANA (Questões Prova)Dokumen6 halamanGENÉTICA HUMANA (Questões Prova)Marcos Felipe TrevisanBelum ada peringkat
- Os 8 Resumos de Genética para Ler em Um Doc SóDokumen90 halamanOs 8 Resumos de Genética para Ler em Um Doc SóVictor da Silva CordeiroBelum ada peringkat
- A Replicação Semiconservativa Do DNADokumen4 halamanA Replicação Semiconservativa Do DNAcarmengatoiBelum ada peringkat
- Mecanismo Molecular Da Replicação Do DNA (Artigo) - Khan AcademyDokumen12 halamanMecanismo Molecular Da Replicação Do DNA (Artigo) - Khan AcademyMariahBelum ada peringkat
- Resumo - 11º Ano - Replicação Do ADNDokumen1 halamanResumo - 11º Ano - Replicação Do ADNHugo Martins100% (1)
- GeneticaDokumen4 halamanGeneticaLuzia ReisBelum ada peringkat
- Aula 6 Material de Apoio Transcrição e TraduçãoDokumen10 halamanAula 6 Material de Apoio Transcrição e TraduçãoRenaly AraujoBelum ada peringkat
- 3º RESUMO BMC, Reparo, Transcrição e Tradução PDFDokumen40 halaman3º RESUMO BMC, Reparo, Transcrição e Tradução PDFrsantanams100% (1)
- Questões de Fixação - Replicação Do DNADokumen3 halamanQuestões de Fixação - Replicação Do DNAMarcos LopesBelum ada peringkat
- Replicação Do DNADokumen5 halamanReplicação Do DNALarissa Lima LiquiBelum ada peringkat
- DNA e Replicação Do DNADokumen4 halamanDNA e Replicação Do DNAGabriel Ferreira100% (1)
- Replicação BidirecionalDokumen7 halamanReplicação BidirecionalAna Amélia Palomino100% (2)
- Continuação Da Lista de Exercícios 1Dokumen3 halamanContinuação Da Lista de Exercícios 1adrianegs50% (2)
- DNA Genes e ReplicaçãoDokumen71 halamanDNA Genes e ReplicaçãoIgor SavoldiBelum ada peringkat
- Cap 3 S - Sol METABOLISMO DO DNA - AlunosDokumen8 halamanCap 3 S - Sol METABOLISMO DO DNA - AlunosGisela SilvaBelum ada peringkat
- Prova 2 de GmolDokumen6 halamanProva 2 de GmolMatheus FelipeBelum ada peringkat
- AULA Replicao2022Dokumen42 halamanAULA Replicao2022Kragen RathmoreBelum ada peringkat
- Bio Aula 2 DNADokumen3 halamanBio Aula 2 DNAMatheus PadilhaBelum ada peringkat
- Perguntas de Treino RespondidasDokumen37 halamanPerguntas de Treino RespondidasseavidafosseBelum ada peringkat
- Documento 32Dokumen7 halamanDocumento 32Ana BeatrizBelum ada peringkat
- Replicação - O RNA Iniciador Da Síntese de DNA - Biologia MolecularDokumen3 halamanReplicação - O RNA Iniciador Da Síntese de DNA - Biologia MolecularFrancivan SantosBelum ada peringkat
- Qa 2Dokumen3 halamanQa 2santiagocordeirosilvaBelum ada peringkat
- Biologia 1Dokumen18 halamanBiologia 1sisojcoelhoBelum ada peringkat
- Replica Ç ÃoDokumen36 halamanReplica Ç ÃoFabricia PredesBelum ada peringkat
- Resumo 2 de Microbio - 230819 - 112306 - 230820 - 222631Dokumen27 halamanResumo 2 de Microbio - 230819 - 112306 - 230820 - 222631Maria Izadora Oliveira CardosoBelum ada peringkat
- M1P1Dokumen9 halamanM1P1Hemily RodriguesBelum ada peringkat
- ResuminhosDokumen9 halamanResuminhosGuilherme GomesBelum ada peringkat
- Duplicação Transcrição e TraduçãoDokumen11 halamanDuplicação Transcrição e TraduçãoSergio Miguel da SilvaBelum ada peringkat
- Introdução Replicação Do DnaDokumen7 halamanIntrodução Replicação Do Dnaarturpanda panzoBelum ada peringkat
- Bruno. Transcrissao e Processamento de DnaDokumen3 halamanBruno. Transcrissao e Processamento de DnaBruno alberto AndreBelum ada peringkat
- Aula T8 - Estrutura Do DNA e ReplicaoDokumen26 halamanAula T8 - Estrutura Do DNA e ReplicaolucianasousabarrospBelum ada peringkat
- Material Genético e Fluxo Da Informação GenéticaDokumen9 halamanMaterial Genético e Fluxo Da Informação Genéticamatheusfed2Belum ada peringkat
- Estrutura e Replicação Do DNADokumen10 halamanEstrutura e Replicação Do DNAGiovanna GoldnerBelum ada peringkat
- Replicação Do DnaDokumen2 halamanReplicação Do Dnacadsus susBelum ada peringkat
- Ficha Replicacao Do DnaDokumen5 halamanFicha Replicacao Do DnacrisvitBelum ada peringkat
- TópicosDokumen45 halamanTópicosCamilaBelum ada peringkat
- A1 Dna - Estrutura e ReplicaçãoDokumen62 halamanA1 Dna - Estrutura e ReplicaçãoPix de ThaynáBelum ada peringkat
- Natureza Química Dos Ácidos NucleicosDokumen9 halamanNatureza Química Dos Ácidos NucleicosnewtonBelum ada peringkat
- Replicação Do DNADokumen4 halamanReplicação Do DNAfradedaiBelum ada peringkat
- Processos Biologicos - Capitulo 4Dokumen57 halamanProcessos Biologicos - Capitulo 4Fiscal PelotenseBelum ada peringkat
- ReplicaçãoDokumen3 halamanReplicaçãoGLEIDSON GUILHERME CARVALHO DA SILVABelum ada peringkat
- Resumo Biologia Molecular AV1: Polímeros. São PolímerosDokumen17 halamanResumo Biologia Molecular AV1: Polímeros. São PolímerosEduarda QueirozBelum ada peringkat
- A Replicação e Transcrição Do Dna - Ext. 2023Dokumen45 halamanA Replicação e Transcrição Do Dna - Ext. 2023Carol Bakos100% (1)
- Replicação, Transcrição e TraduçãoDokumen37 halamanReplicação, Transcrição e TraduçãoNatalia Maria MilhoratiBelum ada peringkat
- DNA e Síntese Protéica PDFDokumen6 halamanDNA e Síntese Protéica PDFMarcela TimossiBelum ada peringkat
- Estudo Dirigido ReplicaçãoDokumen6 halamanEstudo Dirigido ReplicaçãoadrianojosedeoliveiraBelum ada peringkat
- Lista Replicação DNADokumen3 halamanLista Replicação DNALucas RodriguesBelum ada peringkat
- Apostila Síntese de PTNS 2Dokumen9 halamanApostila Síntese de PTNS 2Ana Clara Campos DuarteBelum ada peringkat
- QUESTIONÁRIO 1 Genetica Molecular Arnaldo AcioleDokumen5 halamanQUESTIONÁRIO 1 Genetica Molecular Arnaldo AcioleAllan DavidBelum ada peringkat
- Resumo Estrutura Do DNA, Replicação, Transcrição, Tradução e MutaçãoDokumen15 halamanResumo Estrutura Do DNA, Replicação, Transcrição, Tradução e MutaçãombtbotinhaBelum ada peringkat
- RESUMO - AV2 GenéticaDokumen8 halamanRESUMO - AV2 GenéticaMarina VieiraBelum ada peringkat
- Aula 3 - Ciclo Celular e Replicação Do DNADokumen62 halamanAula 3 - Ciclo Celular e Replicação Do DNAFrancielli Valerio de OliveiraBelum ada peringkat
- Trabo A Ser Feito 2024Dokumen8 halamanTrabo A Ser Feito 2024Adriano VictorBelum ada peringkat
- Conteúdo Do TrabalhoDokumen11 halamanConteúdo Do Trabalhojoaodalasjw26Belum ada peringkat
- Lentes OftálmicasDokumen4 halamanLentes OftálmicasE.e. Raposo TavaresBelum ada peringkat
- Saúde e Segurança No Trabalho - Unidades de Alimentação e NutriçãoDokumen23 halamanSaúde e Segurança No Trabalho - Unidades de Alimentação e NutriçãoGelci Ribeiro75% (4)
- Farmacologia AvançadaDokumen3 halamanFarmacologia AvançadaAlex MartinsBelum ada peringkat
- Aula Cólica RoselmaDokumen42 halamanAula Cólica Roselmakr6ggnrn8cBelum ada peringkat
- Leobel Pompa MorenoDokumen41 halamanLeobel Pompa MorenotharcyllamedBelum ada peringkat
- Manual Diu 08 2018 PDFDokumen72 halamanManual Diu 08 2018 PDFIvana FreitasBelum ada peringkat
- Chas para Queimar GorduraDokumen17 halamanChas para Queimar GorduraAntonio Roque Cardoso100% (1)
- Prótese Parcial FixaDokumen19 halamanPrótese Parcial FixaLanerBotrelBelum ada peringkat
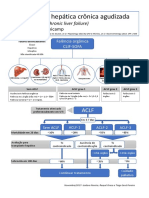
- ACLF - Protocolo UnicampDokumen1 halamanACLF - Protocolo UnicampRubia Moresi Vianna De OliveiraBelum ada peringkat
- Síndrome de Asperger - Aspectos Científicos e EducacionaisDokumen14 halamanSíndrome de Asperger - Aspectos Científicos e EducacionaisconpesBelum ada peringkat
- Morte Encefálica-2Dokumen39 halamanMorte Encefálica-2Celso Joao Mauer100% (1)
- Doença Hipertensiva Específica Da Gestação (DHEG)Dokumen28 halamanDoença Hipertensiva Específica Da Gestação (DHEG)Kelvin InácioBelum ada peringkat
- Oleo Groselha Negra Laszlo PDFDokumen4 halamanOleo Groselha Negra Laszlo PDFRossana LuminaBelum ada peringkat
- Aula+5 Microbiota+VaginalDokumen16 halamanAula+5 Microbiota+VaginalAgna TavaresBelum ada peringkat
- Cartilha Prevencao QuedasDokumen11 halamanCartilha Prevencao Quedasjrjuliao100% (1)
- Questionário ProustDokumen3 halamanQuestionário ProustOlavo MoreiraBelum ada peringkat
- Johrei Pontos VitaisDokumen41 halamanJohrei Pontos VitaisHenri Daniel Lalli100% (10)
- ME Claudia Bonilla Original PDFDokumen143 halamanME Claudia Bonilla Original PDFMax MaxBelum ada peringkat
- Medidas de Prevenção de Infecções Relacionada À Assistência À SaúdeDokumen50 halamanMedidas de Prevenção de Infecções Relacionada À Assistência À SaúdeDaniele SoaresBelum ada peringkat
- Sistema Respiratório QuestõesDokumen3 halamanSistema Respiratório Questõesgillianlelis2Belum ada peringkat
- Conhecendo A Rede de ReabilitacaoDokumen32 halamanConhecendo A Rede de Reabilitacaoscully.xa51Belum ada peringkat
- LESÕES BENIGNAS DA MAMA InternatoDokumen31 halamanLESÕES BENIGNAS DA MAMA InternatoFábio Brito BragaBelum ada peringkat
- Primeiros Socorros em Idosos PDFDokumen22 halamanPrimeiros Socorros em Idosos PDFMónica AlmeidaBelum ada peringkat
- Dieta de South BeachDokumen2 halamanDieta de South BeachAlexsandro AndradeBelum ada peringkat
- Coleção SENAR 236 Bovino Primeiros SocorrosDokumen74 halamanColeção SENAR 236 Bovino Primeiros SocorrosAdimar Cardoso JúniorBelum ada peringkat
- Documento Questionario CC e CMEDokumen10 halamanDocumento Questionario CC e CMEedna647Belum ada peringkat
- Aumente A Sua Massa Muscular e o Seu TamanhoDokumen12 halamanAumente A Sua Massa Muscular e o Seu TamanhoMarcus NeivaBelum ada peringkat
- Novo Protocolo de PodologiaDokumen11 halamanNovo Protocolo de PodologiaPedro Corrêa100% (3)
- Prova UVV 2018-1Dokumen29 halamanProva UVV 2018-1Bruno RebequiBelum ada peringkat
- Leguminosas 2016 PARTILHARDokumen128 halamanLeguminosas 2016 PARTILHARLippy Manuel0% (1)