Anda mungkin juga menyukai
- Practica N 06Dokumen8 halamanPractica N 06Maicol RojasBelum ada peringkat
- Linfopoyesis PDFDokumen92 halamanLinfopoyesis PDFjuanv48Belum ada peringkat
- Prueba Quinto BasicoDokumen4 halamanPrueba Quinto BasicoJavi FernandaBelum ada peringkat
- Caso Clínico 3 Control de MovimientoDokumen3 halamanCaso Clínico 3 Control de MovimientoAndrea Lucía Rodríguez100% (1)
- Caso Clinico ...Dokumen6 halamanCaso Clinico ...Jesus GuerreroBelum ada peringkat
- Acidos Carboxilicos ExpoDokumen17 halamanAcidos Carboxilicos ExpoJudith VilaBelum ada peringkat
- Extraccion de Lipidos de Cerebro Brain LDokumen4 halamanExtraccion de Lipidos de Cerebro Brain LJosue AlvarezBelum ada peringkat
- Fármacos Agonistas CatecolaminergicosDokumen18 halamanFármacos Agonistas CatecolaminergicosMartina Alarcon AcevedoBelum ada peringkat
- Embriología Del Sistema CardiovascularDokumen11 halamanEmbriología Del Sistema CardiovascularEduardoBelum ada peringkat
- Clase 4. Aminoacidos y ProteinasDokumen60 halamanClase 4. Aminoacidos y Proteinasdaniel45Belum ada peringkat
- AsparaginaDokumen3 halamanAsparaginaMaribel PerezBelum ada peringkat
- Práctica 7. Cuantificación de TriglicéridosDokumen7 halamanPráctica 7. Cuantificación de TriglicéridosJesús SalazarBelum ada peringkat
- EsporulaciónDokumen1 halamanEsporulaciónYizua Gomz100% (1)
- Puntos de RestricciónDokumen20 halamanPuntos de RestricciónLuis Copalcua LopantziBelum ada peringkat
- Scheznarda EndocrinoDokumen7 halamanScheznarda Endocrinoricardo_roldan_36Belum ada peringkat
- Aspartato ALEDokumen14 halamanAspartato ALESolanny BenítezBelum ada peringkat
- Fosforilación OxidativaDokumen32 halamanFosforilación OxidativaJasonRodriguezGuerraBelum ada peringkat
- Difusion y Medio Ambiente InternoDokumen25 halamanDifusion y Medio Ambiente InternoAbner Chavez100% (2)
- Cuestionario Metabolismo IntermediarioDokumen8 halamanCuestionario Metabolismo IntermediarioAmyEBermudezQuero0% (1)
- Sintesis y Oxidacion de Nucleotidos-1Dokumen31 halamanSintesis y Oxidacion de Nucleotidos-1Rony Torres ValleBelum ada peringkat
- Grupo 11 Tejido Hematopoyético PreguntasDokumen2 halamanGrupo 11 Tejido Hematopoyético PreguntasMateoo06Belum ada peringkat
- Metabolismo de Acidos Grasos.Dokumen7 halamanMetabolismo de Acidos Grasos.Mariel AybarBelum ada peringkat
- Tecnicas CitogeneticasDokumen24 halamanTecnicas CitogeneticasluisBelum ada peringkat
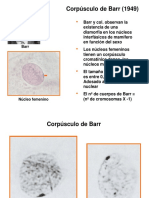
- Corpúsculo de BarrDokumen20 halamanCorpúsculo de BarrNoemi Chura0% (1)
- Complejo de Ataque A La MembranaDokumen9 halamanComplejo de Ataque A La MembranaRoward RojasBelum ada peringkat
- Actividad de Las GrasasDokumen4 halamanActividad de Las Grasasclaudia marcela rubio rojasBelum ada peringkat
- Enfermedades MonogénicasDokumen31 halamanEnfermedades MonogénicasDiabloBelum ada peringkat
- Proteinas de CoagulacionDokumen31 halamanProteinas de CoagulacionHematoped 20 novBelum ada peringkat
- SINTESIS DE COLESTEROL ResumenDokumen4 halamanSINTESIS DE COLESTEROL ResumenXena Katherine Gutarra SosaBelum ada peringkat
- Bacterias PrototrofasDokumen1 halamanBacterias PrototrofasJosé David Ronquillo CandoBelum ada peringkat
- GlicoproteinasDokumen45 halamanGlicoproteinasAlexis PomaBelum ada peringkat
- Metabolismo de Los Hidratos de Carboo y Formacion Del Trifosfato de AdenosinaDokumen3 halamanMetabolismo de Los Hidratos de Carboo y Formacion Del Trifosfato de AdenosinaDaysi camila Vilca CoaBelum ada peringkat
- Acdos Nucleicos ProyectoDokumen5 halamanAcdos Nucleicos ProyectoJosePonceVeraBelum ada peringkat
- Enfermedades MitocondrialesDokumen15 halamanEnfermedades MitocondrialesJuan Domingo Davila GilBelum ada peringkat
- Genética 7Dokumen16 halamanGenética 7axel floresBelum ada peringkat
- MORALES RENTERIA AILYN - Practicas DigestivoDokumen9 halamanMORALES RENTERIA AILYN - Practicas DigestivoArturo TejedaBelum ada peringkat
- Fosforilación Oxidativa - ResumenDokumen10 halamanFosforilación Oxidativa - ResumenKevinDelaCruzBelum ada peringkat
- Cuestionario de Glucidos y MetabolismoDokumen5 halamanCuestionario de Glucidos y MetabolismoIAGO SAMUEL DE SOUZA REISBelum ada peringkat
- INTRODUCCIÓNDokumen54 halamanINTRODUCCIÓNSergio RamosBelum ada peringkat
- Epitelial Mapa ConceptualDokumen18 halamanEpitelial Mapa ConceptualmichaelBelum ada peringkat
- 3CM3, AGWP, Reporte Practica No.5Dokumen15 halaman3CM3, AGWP, Reporte Practica No.5Wendy AguileraBelum ada peringkat
- (Inmunologia) Resumen Teorico 14Dokumen3 halaman(Inmunologia) Resumen Teorico 14Samir Torres ChauBelum ada peringkat
- Operón TriptófanoDokumen6 halamanOperón Triptófano1735171012Belum ada peringkat
- Cuestionario Transduccion de SeñalesDokumen2 halamanCuestionario Transduccion de SeñalesVictor Lecca50% (2)
- Receptor Nicotinico ACh PDFDokumen7 halamanReceptor Nicotinico ACh PDFlabavillaBelum ada peringkat
- Agonistas ColinérgicosDokumen13 halamanAgonistas ColinérgicosruloBelum ada peringkat
- Vias de Inoculacion ParascribDokumen3 halamanVias de Inoculacion ParascribReinilda SaldivarBelum ada peringkat
- Cromosomas y Cariotipo HumanoDokumen6 halamanCromosomas y Cariotipo HumanoMARIA DE LOS ANGELES MARIMON ESTEVESBelum ada peringkat
- AmilasaDokumen12 halamanAmilasaPaola Borlone ValderramaBelum ada peringkat
- 1.introduccion Al MetabolismoDokumen9 halaman1.introduccion Al MetabolismoAndres Felipe Prieto AlarconBelum ada peringkat
- Presentacion BiofisicaDokumen1 halamanPresentacion BiofisicaAngelica Stephania Ramirez Gracia100% (2)
- Regulacion de La Exprecion Genica, Sistema InmunitarioDokumen17 halamanRegulacion de La Exprecion Genica, Sistema InmunitarioKleverxithoDiazCiezaBelum ada peringkat
- Corazón, EpónimosDokumen2 halamanCorazón, EpónimosFrank RitikonBelum ada peringkat
- Aminoacidos y Peptidos-Harper. Bioquimica Ilustrada 28thDokumen7 halamanAminoacidos y Peptidos-Harper. Bioquimica Ilustrada 28thjhonny gonzalezBelum ada peringkat
- El HigadoDokumen40 halamanEl HigadoJenny Jazmine Perez SandovalBelum ada peringkat
- Desarrollo Del Aparato RespiratorioDokumen2 halamanDesarrollo Del Aparato RespiratorioAngel Stevens Garcia CallaoBelum ada peringkat
- Genes y Control de La Expresión GenéticaDokumen12 halamanGenes y Control de La Expresión GenéticaYAHIZIRI JANAIL VENTURA SAMAYOABelum ada peringkat
- Regulación de La Expresión Génica en ProcariontesDokumen32 halamanRegulación de La Expresión Génica en ProcariontesBenjamin Padilla ParodiBelum ada peringkat
- Regulacion Genica en ProcariotasDokumen10 halamanRegulacion Genica en ProcariotasMaría Juliana Rojas MoncadaBelum ada peringkat
- Regulación GénicaDokumen40 halamanRegulación GénicaISBELIAC7Belum ada peringkat
- 2.5 OperonesDokumen26 halaman2.5 OperonesAlonso ZavalaBelum ada peringkat
- SR Planif Fliar Misoprostol UsoDokumen4 halamanSR Planif Fliar Misoprostol UsoRuth Esperanza Ferreyra CayoBelum ada peringkat
- EC Calidad y ProductividadDokumen4 halamanEC Calidad y ProductividadKristian CobaBelum ada peringkat
- Jose A Santamaria O: Total $469.71Dokumen2 halamanJose A Santamaria O: Total $469.71Kristian CobaBelum ada peringkat
- EC Calidad y ProductividadDokumen4 halamanEC Calidad y ProductividadKristian CobaBelum ada peringkat
- Jose A Santamaria O: Total $469.71Dokumen2 halamanJose A Santamaria O: Total $469.71Kristian CobaBelum ada peringkat
- SR Planif Fliar Misoprostol UsoDokumen4 halamanSR Planif Fliar Misoprostol UsoRuth Esperanza Ferreyra CayoBelum ada peringkat
- Tarea LaboratoriosDokumen23 halamanTarea LaboratoriosKristian CobaBelum ada peringkat
- Los Cuerpos Geométricos Están Por Todos LadosDokumen1 halamanLos Cuerpos Geométricos Están Por Todos LadosKristian CobaBelum ada peringkat
- Canciones KevinDokumen3 halamanCanciones KevinKristian CobaBelum ada peringkat
- Como Prevenirla ContaminacionDokumen2 halamanComo Prevenirla ContaminacionKristian CobaBelum ada peringkat
- Noticia Nacional MiDokumen6 halamanNoticia Nacional MiKristian CobaBelum ada peringkat
- Mecanismos Postraduccionales Unidad 4Dokumen21 halamanMecanismos Postraduccionales Unidad 4Kristian CobaBelum ada peringkat
- Hugo Cesion de Derechos Tio MiguelDokumen1 halamanHugo Cesion de Derechos Tio MiguelKristian CobaBelum ada peringkat
- Copia de Plan de Trabajo CircuitosDokumen1 halamanCopia de Plan de Trabajo CircuitosKristian CobaBelum ada peringkat
- 26Dokumen6 halaman26Kristian CobaBelum ada peringkat
- Mapa ConceptualDokumen2 halamanMapa ConceptualKristian CobaBelum ada peringkat
- Sistema Segundo OrdennnnDokumen4 halamanSistema Segundo OrdennnnKristian CobaBelum ada peringkat
- El Planeta Se Está CalentandoDokumen2 halamanEl Planeta Se Está CalentandoKristian CobaBelum ada peringkat
- Examn de Bioquimica Trascripcion (Uinidad 5)Dokumen2 halamanExamn de Bioquimica Trascripcion (Uinidad 5)Kristian CobaBelum ada peringkat
- Examn Unidad 6 (BIOQUIMICA)Dokumen1 halamanExamn Unidad 6 (BIOQUIMICA)Kristian CobaBelum ada peringkat
- Queremos Que Esta ReuniónDokumen5 halamanQueremos Que Esta ReuniónKristian CobaBelum ada peringkat
- Diagramas de BloquesDokumen11 halamanDiagramas de BloquesKristian CobaBelum ada peringkat
- Actividad 7 CDokumen2 halamanActividad 7 CKristian CobaBelum ada peringkat
- AdsorcionDokumen2 halamanAdsorcionmako_dbzBelum ada peringkat
- Adjunto EducDokumen6 halamanAdjunto EducKristian CobaBelum ada peringkat
- Actividad 6Dokumen2 halamanActividad 6Kristian CobaBelum ada peringkat
- Unidad 6Dokumen12 halamanUnidad 6Asahel Soto GarciaBelum ada peringkat
- Adelita Del Carmen Silvaran MartinezDokumen1 halamanAdelita Del Carmen Silvaran MartinezKristian CobaBelum ada peringkat
- Actividad 7-b Ensayo de La Evolucion de La Carrera de Ingenieria AmbientalDokumen3 halamanActividad 7-b Ensayo de La Evolucion de La Carrera de Ingenieria AmbientalKristian CobaBelum ada peringkat
- Resumen Receptores Acoplados A Proteínas G y Sus Múltiples FacetasDokumen2 halamanResumen Receptores Acoplados A Proteínas G y Sus Múltiples FacetasMARTHA CRUZBelum ada peringkat
- 001 Introducion A Diana BiologicasDokumen27 halaman001 Introducion A Diana BiologicasEstefany Licet Miranda ArriagaBelum ada peringkat
- Biofísica - La TorreDokumen27 halamanBiofísica - La TorreCINDY YAQUELIN IÑIGUA CUROBelum ada peringkat
- Mov Biologicos MMNQHDokumen19 halamanMov Biologicos MMNQHRene vasquez telloBelum ada peringkat
- Inhibidores de La ProteasaDokumen2 halamanInhibidores de La ProteasaClaudia PeñaBelum ada peringkat
- Dialnet EnBuscaDeLaEstructuraDelADN 719046Dokumen11 halamanDialnet EnBuscaDeLaEstructuraDelADN 719046lucasBelum ada peringkat
- VIH - SIDA PDF Por MediversiaDokumen38 halamanVIH - SIDA PDF Por MediversiaKELLY JOHANNA BOLAÑOS PAYABelum ada peringkat
- 5 Variaciones de Las Leyes de MendelDokumen44 halaman5 Variaciones de Las Leyes de MendelbuzztintinBelum ada peringkat
- Ácidos NucleicosDokumen5 halamanÁcidos NucleicosEduardo Isidoro ReyesBelum ada peringkat
- Formas PrecelularesDokumen1 halamanFormas PrecelularesLu borjasBelum ada peringkat
- 06 Fundamentos de EmbriologíaDokumen11 halaman06 Fundamentos de EmbriologíaSimón CisnerosBelum ada peringkat
- Farmacotecnia I para FinalDokumen50 halamanFarmacotecnia I para FinalAriel AcostaBelum ada peringkat
- P7 - Ingeniería de Proteínas - Sánchez Zamorano Julio CésarDokumen10 halamanP7 - Ingeniería de Proteínas - Sánchez Zamorano Julio CésarJulio César Sánchez ZamoranoBelum ada peringkat
- Desarrollo de Los Linfocitos B ResumenDokumen12 halamanDesarrollo de Los Linfocitos B ResumenJGRBBKDVRBelum ada peringkat
- Mn-Epd-01 Manual de Bioseguridad-20182Dokumen157 halamanMn-Epd-01 Manual de Bioseguridad-20182abigailBelum ada peringkat
- Practica 6Dokumen3 halamanPractica 6Jonathan RaxcacóBelum ada peringkat
- NEURONADokumen3 halamanNEURONAAdriel Galvan RochaBelum ada peringkat
- Crucigrama FacilDokumen1 halamanCrucigrama FacilFernando RodríguezBelum ada peringkat
- Trabjo de BiologíaDokumen11 halamanTrabjo de Biologíatonixd123456789Belum ada peringkat
- El Amarrillo Martius Es Un Colorante Que Se Prepara Cuando ElDokumen4 halamanEl Amarrillo Martius Es Un Colorante Que Se Prepara Cuando ElfernandoluisBelum ada peringkat
- Trabajo de E. ColiDokumen46 halamanTrabajo de E. ColiMagglis Zerpa AvelloBelum ada peringkat
- Proyecto de Biologia 2do de Bachillerato Parcial 2Dokumen18 halamanProyecto de Biologia 2do de Bachillerato Parcial 2Yury RamirezBelum ada peringkat
- Transaminación y Desaminación de ProteínasDokumen24 halamanTransaminación y Desaminación de ProteínasCITLALLI MU�OZ ANDRADEBelum ada peringkat
- Ciclo KerbsDokumen3 halamanCiclo KerbsAllan Kun DamasBelum ada peringkat
- El Reino Monera - PracticaDokumen6 halamanEl Reino Monera - PracticaMario Salinas100% (1)
- Presentación Ciencia Biología para Principiantes Células Orgánico Ilustrado Morado OscuroDokumen8 halamanPresentación Ciencia Biología para Principiantes Células Orgánico Ilustrado Morado OscuroFranklin Omonte SanchezBelum ada peringkat
- Evolucion MicrobianaDokumen10 halamanEvolucion MicrobianaRonny Peña BáezBelum ada peringkat
- CromosomaDokumen4 halamanCromosomalorenyis maitaBelum ada peringkat
- Examen de Subsanación Cuarto AñoDokumen2 halamanExamen de Subsanación Cuarto AñoCarlos Maguiña100% (1)