Anda mungkin juga menyukai
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Ce Aduce Nou Interpretarea PolisomnografieDokumen7 halamanCe Aduce Nou Interpretarea PolisomnografieLaura MoiseBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
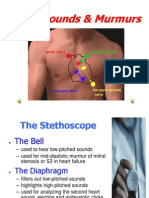
- Heart Sounds and MurmursDokumen38 halamanHeart Sounds and MurmursLaura Moise100% (5)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- A Guide To Performing Quality Assured Diagnostic SpirometryDokumen28 halamanA Guide To Performing Quality Assured Diagnostic SpirometryLaura MoiseBelum ada peringkat
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- ATS ERS Spirometry Standards 2005Dokumen20 halamanATS ERS Spirometry Standards 2005Laura MoiseBelum ada peringkat
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- Diabetes Alphabet Strategy SymbolsDokumen9 halamanDiabetes Alphabet Strategy SymbolsLaura MoiseBelum ada peringkat
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Bajaj Holdings & Investment Ltd. - Research Center: Balance SheetDokumen6 halamanBajaj Holdings & Investment Ltd. - Research Center: Balance Sheetsarathkumarreddy855081Belum ada peringkat
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Region XiDokumen2 halamanRegion XiGarcia, Rafael Rico O.Belum ada peringkat
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- SSP 861603 - EN - Tire Pressure Monitoring SystemsDokumen42 halamanSSP 861603 - EN - Tire Pressure Monitoring Systemsa.diedrichsBelum ada peringkat
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- SAP Logistics Process ScopeDokumen3 halamanSAP Logistics Process ScopecasiloBelum ada peringkat
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- This Study Resource WasDokumen2 halamanThis Study Resource Waskaye nicolasBelum ada peringkat
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Sistem Informasi Pencatatan Data Pelanggan Pada Telkomsel Grapari Banda AcehDokumen6 halamanSistem Informasi Pencatatan Data Pelanggan Pada Telkomsel Grapari Banda AcehJurnal JTIK (Jurnal Teknologi Informasi dan Komunikasi)Belum ada peringkat
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- 20 Heirs of Alfredo Bautista VS LindoDokumen3 halaman20 Heirs of Alfredo Bautista VS LindoJerome LeañoBelum ada peringkat
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Interview With Hohepa Mapiria Joseph - Joe - Murphy Royal Regent 7 July 2003Dokumen61 halamanInterview With Hohepa Mapiria Joseph - Joe - Murphy Royal Regent 7 July 2003kiwiit100% (8)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- Nabard Working CapitalDokumen12 halamanNabard Working CapitalJaspreet SinghBelum ada peringkat
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- PMAIZMTUSDMDokumen6 halamanPMAIZMTUSDMLinh TranBelum ada peringkat
- 10 Rules of Statcon by Atty Marcus NeelyDokumen4 halaman10 Rules of Statcon by Atty Marcus NeelyMorin OcoBelum ada peringkat
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Assignment Practice ProblemsDokumen5 halamanAssignment Practice ProblemsibrahimmoizBelum ada peringkat
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- Ksiidc and KssidcDokumen13 halamanKsiidc and KssidckasperBelum ada peringkat
- Qualitative Analysis of Selected Medicinal Plants, Tamilnadu, IndiaDokumen3 halamanQualitative Analysis of Selected Medicinal Plants, Tamilnadu, IndiaDiana RaieBelum ada peringkat
- Kids Cooking Teams: That's FreshDokumen74 halamanKids Cooking Teams: That's FreshNCB School of Herbalism & Holistic HealthBelum ada peringkat
- Individual Ability in SoftwareDokumen9 halamanIndividual Ability in Softwaredhana0809100% (4)
- Wipro Home Office RangeDokumen8 halamanWipro Home Office RangePrashant RawatBelum ada peringkat
- RagragsakanDokumen6 halamanRagragsakanRazel Hijastro86% (7)
- Literature Review Electrical EngineeringDokumen6 halamanLiterature Review Electrical Engineeringtkpmzasif100% (1)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- Pico Power: A Boon For Rural Electrification: Rajat KapoorDokumen8 halamanPico Power: A Boon For Rural Electrification: Rajat KapoorBamBelum ada peringkat
- Tamang GrammarDokumen12 halamanTamang Grammarsoftdina100% (1)
- Clause: Extra Element + Independent Clause Dependent ClauseDokumen1 halamanClause: Extra Element + Independent Clause Dependent ClauseTieng HuangBelum ada peringkat
- Es PS 0614Dokumen6 halamanEs PS 0614陳相如Belum ada peringkat
- Fractal Blaster Trading Strategy ReportDokumen22 halamanFractal Blaster Trading Strategy ReportIcky IckyBelum ada peringkat
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- YazdiDokumen7 halamanYazditrs1970Belum ada peringkat
- Admission Sos 2013-14090513 PDFDokumen21 halamanAdmission Sos 2013-14090513 PDFmanoj31285manojBelum ada peringkat
- Apy1 Kinematics Review - 2015 - KeyDokumen13 halamanApy1 Kinematics Review - 2015 - KeyemadBelum ada peringkat
- Founders' ShareDokumen2 halamanFounders' ShareHenry BerlangaBelum ada peringkat
- 2017 - Behavioral Emergencies - Geriatric Psychiatric PatientDokumen14 halaman2017 - Behavioral Emergencies - Geriatric Psychiatric PatientAna María Arenas DávilaBelum ada peringkat
- B2 UNIT 4 Test StandardDokumen6 halamanB2 UNIT 4 Test StandardВладимир РанцовBelum ada peringkat
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)