Anda mungkin juga menyukai
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
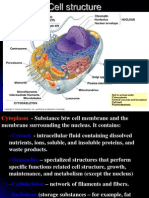
- Cell CytoplasmDokumen55 halamanCell Cytoplasmnicolaer111100% (1)
- Unit 1 Anatomy, Physiology and The Human Body: Learning OutcomesDokumen91 halamanUnit 1 Anatomy, Physiology and The Human Body: Learning OutcomesDeolita BadiangBelum ada peringkat
- Gesc 107Dokumen12 halamanGesc 107Nippu SharmaBelum ada peringkat
- ICSE QUESTION PAPER Biology 2010 Class 10 PDFDokumen11 halamanICSE QUESTION PAPER Biology 2010 Class 10 PDFLokesh MalikBelum ada peringkat
- IB Biology HL: 11.1 Antibody Production and VaccinationDokumen7 halamanIB Biology HL: 11.1 Antibody Production and VaccinationpetraBelum ada peringkat
- Skeletal System4Dokumen1 halamanSkeletal System4Elisha DienteBelum ada peringkat
- Cervical and Shoulder 2020Dokumen14 halamanCervical and Shoulder 2020Eduardo OlivaBelum ada peringkat
- Chapter 12: The Back: Vertebral ColumnDokumen7 halamanChapter 12: The Back: Vertebral ColumnJyrra NeriBelum ada peringkat
- Development of The Fetal Membranes and PlacentaDokumen4 halamanDevelopment of The Fetal Membranes and PlacentaBandula KusumsiriBelum ada peringkat
- PMDC NEB Exams Free Course BookletDokumen8 halamanPMDC NEB Exams Free Course BookletDaxtren HicksBelum ada peringkat
- Rujukan OktoberDokumen15 halamanRujukan Oktoberendang sunarmBelum ada peringkat
- Patient Based PathophysiologyDokumen2 halamanPatient Based PathophysiologyDeinielle Magdangal RomeroBelum ada peringkat
- EMB5, PDFDokumen7 halamanEMB5, PDFSafura IjazBelum ada peringkat
- Pigeon DissectionDokumen10 halamanPigeon DissectionAnonymous 3NmUUDtBelum ada peringkat
- VI 8 Body MovementsDokumen16 halamanVI 8 Body MovementsManisha VarmaBelum ada peringkat
- Connective TissueDokumen62 halamanConnective Tissuehnm mnhBelum ada peringkat
- Fate Mapping - Gastrulation - Lec Notes - KMZRDokumen27 halamanFate Mapping - Gastrulation - Lec Notes - KMZRSaad SakalBelum ada peringkat
- The Functional Anatomy of PhonationDokumen15 halamanThe Functional Anatomy of PhonationCarolyn BensonBelum ada peringkat
- Ai Chi - Meridians2Dokumen4 halamanAi Chi - Meridians2sale18100% (1)
- Torehj 6 1 PDFDokumen20 halamanTorehj 6 1 PDFSaifuddin HaswareBelum ada peringkat
- The Final Ultimate CML Case Pres Final 1 1Dokumen113 halamanThe Final Ultimate CML Case Pres Final 1 1Levy Garcia SanchezBelum ada peringkat
- CH 8 Digestive System: StructuresDokumen18 halamanCH 8 Digestive System: StructuresBheru LalBelum ada peringkat
- The Skeletal SystemDokumen3 halamanThe Skeletal SystemMiguel GumatayBelum ada peringkat
- Digestive System and Respiratory SystemDokumen10 halamanDigestive System and Respiratory SystemKathMae BoaBelum ada peringkat
- Gram Staining Protocol or ProcedureDokumen4 halamanGram Staining Protocol or Procedurearavindbt2523Belum ada peringkat
- Byjus Com Biology Preparation and Study of Transverse Section of Dicot and Monocot Roots and StemsDokumen1 halamanByjus Com Biology Preparation and Study of Transverse Section of Dicot and Monocot Roots and Stemssmm MamaBelum ada peringkat
- Exchange TransfusionDokumen35 halamanExchange Transfusionsobinjohnpkl100% (2)
- Introduction To The Neurologic ExaminationDokumen38 halamanIntroduction To The Neurologic ExaminationYudhistira YuliandraBelum ada peringkat
- AP Module4 TasksBC-DiazDokumen5 halamanAP Module4 TasksBC-DiazJohn Rafael DiazBelum ada peringkat
- Astrocyte: Structure & FunctionDokumen7 halamanAstrocyte: Structure & Functionمحمود الموسويBelum ada peringkat