Apostila de Nutrição Prof (1) - Walter
Diunggah oleh
Paulo Henrique CavalcanteJudul Asli
Hak Cipta
Format Tersedia
Bagikan dokumen Ini
Apakah menurut Anda dokumen ini bermanfaat?
Apakah konten ini tidak pantas?
Laporkan Dokumen IniHak Cipta:
Format Tersedia
Apostila de Nutrição Prof (1) - Walter
Diunggah oleh
Paulo Henrique CavalcanteHak Cipta:
Format Tersedia
Universidade Federal de Minas Gerais Escola de Veterinria Departamento de Zootecnia
CADERNOS DE NUTRIO DE NO-RUMINANTES
Walter Motta Ferreira1
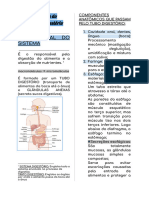
SISTEMA DIGESTIVO DOS NO RUMINANTES. I. INTRODUO. Os animais no podem utilizar diretamente os nutrientes e a energia dos alimentos; atravs dos processos que acontecem no trato gastrointestinal (TGI) que os alimentos so reduzidos a um tal nvel molecular ou a um estado de solubilidade que permitem a absoro de seus nutrientes e a energia. Os processos digestivos supem a diminuio do tamanho das partculas do alimento, a solubilizao de seus componentes qumicos, a hidrlise de suas macro-molculas (polissacardeos, triglicrides e protenas) at molculas de menor peso e complexidade estrutural (acares, cidos graxos, aminocidos), a transformao dos componentes qumicos e a absoro dos nutrientes em sua forma mais simples atravs da parede do (TGI), passando ao sangue ou ao sistema linftico e, por seu intermdio, aos diferentes tecidos e rgos do animal. O objetivo principal destes processos visa o fornecimento das estruturas qumicas necessrias para substituir as j utilizadas e a energia gastada pelo animal nos processos biolgicos destinados a sua mantena como organismo vivo e as empregadas na sntese dos nutrientes (protena, gordura, glicognio) que deposita em seu corpo ou nos produtos que entrega ao homem (ovos, leite) e ao trabalho muscular; assim, por exemplo, o amido do alimento fornecido as porcas em lactao digerido no TGI at glicose que absorvida para ser utilizada em parte como fonte de energia para a realizao de trabalho, na sntese de lactose e gordura do leite e para a sntese de gordura muscular e tecido adiposo destinados acumulao de reservas corporais. O processo digestivo compreende, portanto, trs fases: Formao de nutrientes simples a partir dos componentes de elevado peso molecular e estrutura qumica complexa que esto contidos nos alimentos; Absoro de nutrientes mais simples e da energia atravs das paredes do TGI ao sangue e sistema linftico;
1
Zootecnista, Especialista em Produo Animal, Mestre em Zootecnia, Doutor em Cincia Animal, Professor Adjunto do Departamento de Zootecnia da Universidade Federal de Minas Gerais.
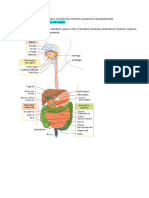
Expulso das fezes, constitudas fundamentalmente pelos resduos no digeridos do alimento e os componentes endgenos do TGI (enzimas secretadas, clulas liberadas e microorganismos). O estudo da digesto nos animais, tanto ruminantes quanto no ruminantes, est associado com os seguintes termos: Apreenso: toma do alimento ou gua; Mastigao: reduo do tamanho da partcula do alimento; Deglutio; Regurgitao: retorno boca do material no digerido; Digesto: clivagem das partculas do alimento em produtos disponveis para serem absorvidos. A digesto pode incluir: a. Foras mecnicas; b. Aes qumicas; c. Atividades enzimticas devidas s enzimas produzidas pelo prprio TGI e pelos microorganismos que o colonizam; 6. Absoro: transferncia de substncias do TGI ao sangue ou ao sistema linftico; 7. Anabolismo: crescimento ou processo de construo de tecidos; 8. Catabolismo: rompimento ou reaes de destruio de nutrientes; 9. Metabolismo: combinao de reaes anablicas e catablicas que acontecem no corpo e liberam energia; 10. Excreo: remoo dos resduos gerados pelos processos digestivos. Com relao ao comportamento alimentar os animais domsticos podem ser divididos Em trs grandes categorias: 1. Herbvoros: espcies animais que so consumidoras de vegetais e dependem totalmente das plantas como fontes de alimento; 2. Carnvoros: espcies animais cuja alimentao est baseada no consumo de carne ou de outros animais; 3. Onvoros: espcies animais que consomem tanto vegetais quanto carne. II. SISTEMA DIGESTIVO DOS NO RUMINANTES. O TGI, s vezes chamado de trato alimentar, representa o espao que vai da boca at o nus ou a cloaca e a travs do qual passa o alimento depois de ser consumido e submetido aos vrios processos digestivos. Os inmeros rgos, glndulas e as outras estruturas envolvidas com o TGI esto associadas com a toma do alimento, mastigao, deglutio e com a digesto e absoro dos nutrientes bem com algumas funes de secreo. O TGI nos no ruminantes apresenta uma estrutura fibrosa muscular coberta por um epitlio que em alguns locais tem-se especializado para a secreo, digesto e a absoro. As paredes do TGI esto constitudas basicamente por quatro membranas ou camadas concntricas, classificadas desde a parte interior exterior da seguinte maneira:
1. Membrana ou camada mucosa.
1. 2. 3. 4. 5.
Camada mais prxima ao lume do TGI, constituda por sua vez por outras trs camadas protegidas por muco produzido por glndulas especializadas:
1.1. Epitlio da superfcie ou lmina epitelial.
Varia entre as diferentes partes do trato sendo freqentemente rico em tecido glandular secretrio.
1.2. Lmina prpria da mucosa ou mucosae.
Consiste de uma camada de tecido conjuntivo que contem inmeros capilares sangneos e, em alguns casos, glndulas mucosas.
1.3. Lmina muscularis mucosae.
Formada por uma ou vrias camadas finas de msculo liso com fibras elsticas, que determinam os movimentos da mucosa sendo enervadas por fibras do sistema simptico.
2. Membrana ou camada submucosa.
Camada de tecido conjuntivo que possui vasos sangneos de dimetro maior e s vezes algumas glndulas. Nela encontra-se o plexo nervoso submucoso com suas clulas ganglionares parasimpticas que inervam as glndulas da mucosa. Este plexo nervoso mais destacado naquelas regies do trato onde a mucosa muito rica em glndulas como por exemplo no estmago, abomaso e proventrculo. 3. Membrana ou camada muscular externa. Camada de tecido muscular responsvel dos movimentos do TGI e constituda, por sua vez, por outras duas camadas:
3.1. Interna, constituda por uma camada de fibras musculares lisas orientadas de
maneira circular. 3.2. Externa, composta de fibras musculares longitudinais. Embora estejam sob a influncia do complexo mientrico prprio, ou sistema nervoso entrico local, achado entre ambas as duas camadas de fibras musculares, os msculos lisos desta membrana recebem a inervao externa atravs das fibras do sistema parassimptico.
4. Membrana ou camada serosa ou adventcia.
uma camada de clulas escamosas simples localizada sobre o tecido conjuntivo mais externo. Normalmente os vasos sangneos e as fibras nervosas percorrem esta membrana antes de ingressar camada muscular. III. CARACTERSTICAS GERAIS DO TRATO GASTROINTESTINAL EM ALGUNS NO RUMINANTES. 1. AVES. BOCA. (1). No tem dentes;
(2). Nas aves a apreenso do alimento responsabilidade principalmente do bico e em menor proporo da lngua. O bico est adaptado para ciscar rapidamente pequenas partculas de alimento, tem uma funo de pina, caso o alimento seja granulado, ou de colher para quando o alimento est sob a forma farelada e serve para reduzir parcialmente o alimento at um tal tamanho que pode ser engolido; (3). A saliva contem amilase e, fundamentalmente, mucina que tem uma funo lubrificante. Nas aves a insalivao muito escassa porquanto suas glndulas salivares esto muito pouco desenvolvidas. A secreo salivar somente de 7 a 25 ml/dia. A baixa secreo de -amilase junto ausncia de mastigao fazem que seja limitada a digesto dos carboidratos na boca. (4). A deglutio do alimento at a faringe tem lugar mediante sucessivos movimentos da cabea para cima e para frente, com a ajuda das papilas existentes na base da lngua as quais esto dirigidas para trs. ESFAGO. relativamente longo, de fcil dilatao e permite a rpida passagem do alimento da boca at o inglvio ou papo (trata-se de uma dilatao do esfago em forma de bolsa membranosa presente em inmeras aves, exceto nas espcies insetvoras) que apresenta as seguintes funes: (1). Serve como um reservatrio para a armazenagem e umedecimento do alimento at que se esvazie ao proventrculo o qual est a continuao do inglvio; (2). Permite a clivagem da amilase salivar; (3). Em algumas espcies acontece a fermentao do amido originando maltose, glicose e cido ltico. PROVENTRCULO (ESTMAGO GLANDULAR). (1). Local de secreo gstrica (produo de HCl e pepsina); pH entre 3,0 e 4,5. A diferena dos monogstricos as aves no produzem lipase gstrica e tanto o HCl quanto a pepsina so produzidos e secretados por um mesmo tipo de clulas; (2). O alimento passa rapidamente por este local (aproximadamente 14 segundos). VENTRCULO (MOELA OU ESTMAGO MUSCULAR). (1). rgo com parede muscular grossa e epitlio cornificado onde reduzido fisicamente o tamanho da partcula do alimento (semelhante mastigao nos mamferos) devido s contraes musculares involuntrias, as quais se apresentam em proporo de uma a cada 20 ou 30 segundos; (2). A parede da moela no tem glndulas de secreo de enzimas, porm est recoberta por uma secreo mucosa espessa; neste local o HCl e a pepsina originados no proventrculo mantm ainda sua atividade; (3). A moela contem normalmente pedras pequenas ou partculas duras que ajudam na moagem das sementes e gros ingeridos; contudo estas no so essenciais para o desenvolvimento da funo de triturao que ali acontece.
INTESTINO DELGADO. (1). Com exceo da lactase vrias das enzimas achadas nos mamferos tambm esto no intestino delgado das aves; (2). O pH do intestino delgado levemente cido; (3). A absoro de nutrientes similar dos mamferos, exceto que nas aves no h secreo da enterogastrona, hormnio que afeta a absoro das gorduras CECO E INTESTINO GROSSO. (1). O TGI das aves tem dois sacos cecos (ceco), no entanto nos mamferos h s um saco; (2). O ceco e intestino grosso so locais de reabsoro dgua; parte da degradao da fibra do alimento e sntese de vitaminas solveis em gua acontece no ceco devido fermentao bacteriana, sendo, contudo, menor estas atividades nas aves quando comparadas com os mamferos, em especial os mamferos herbvoros no ruminantes; (3). O intestino grosso muito curto (5-10 cm) e esvazia o seu contedo dentro da cloaca, cavidade onde saem tambm as vias urogenitais e de donde o material fecal e urinrio pode ser descarregado ao meio. As aves no alojadas em gaiolas ingerem os dejetos depositados no cho; contudo, este fenmeno de pouca importncia e no possui a significncia que tem para coelhos.
2. CO.
BOCA. Os ces devoram o alimento sem mastigar; uma vez que o alimento apreendido e mastigado, estimula-se a produo de saliva devido viso e o cheiro do alimento. A produo de saliva facilita a deglutio do alimento. DEGLUTICO. O alimento transferido da boca at o estmago atravs de um tubo relativamente curto chamado de esfago; embora neste local no sejam produzidas enzimas, as suas clulas adicionam grandes quantidades de muco que facilitam a passagem do alimento. A presena do alimento no esfago estimula o peristaltismo empurrando-o para o estmago. Entre o esfago e o estmago existe a crdia que estimulada pela onda peristltica permitindo a passagem do alimento; a presso gerada no estmago no causa o relaxamento do esfago, sendo, portanto, improvvel a volta do alimento boca, exceto em circunstncias anormais como por exemplo vmito. ESTMAGO.
(1). O estmago nos ces funciona como reservatrio permitindo que o alimento seja ingerido na forma de refeies espaadas ao invs que continuamente. Neste local do TGI onde comena realmente a digesto das protenas; (2). O estmago tambm regula o fluxo de materiais para o intestino delgado. (3). Funcionalmente pode ser dividido em duas pores: Corpo: Possui paredes muito elsticas o que permite a armazenagem de grandes quantidades de alimento sem qualquer aumento da presso intragstrica. A mucosa do corpo do estmago secreta muco. HCl e proteases, sendo que uma delas, a pepsina, produzida em forma inativa como pepsinognio seguindo os mesmos mecanismos de controle hormonal e nervoso que para as outras espcies animais no ruminates. Antro: A mucosa antral, em contraste com a mucosa do corpo, produz uma soluo alcalina pobre em enzimas.
INTESTINO DELGADO. O alimento parcialmente digerido no estmago passa ao dudeno. A velocidade pela qual o estmago libera o quimo (mistura lquida espessa e leitosa formada no estmago entre o alimento e as secrees digestivas) para o duodeno controlada pelo esfincter pilrico. No intestino delgado do ce acontecem os mesmos processos digestivos sobre os nutrientes e a energia que nas outras espcies no ruminantes. INTESTINO GROSSO. (1). No apresenta vilosidades; (2). Sua superfcie de absoro limitada; (3). Embora uma parcela pequena do alimento e da gua ingerido chega at o intestino grosso, as colnias de bactrias residentes neste local do TGI so capacez de digerir parcialmente algumas protenas e resduos dos alimentos fibrosos. 3. COELHO. Anatomicamente o coelho apresenta estmago e ceco bastante desenvolvidos (com capacidade de conter cerca de 80% da digesta) e esto bem adaptados fermentao posterior ao intestino delgado. Outra particularidade presente nesta espcie est associada com a cecotrofia que a capacidade que tem os coelhos de reingerir parte do material fecal, as denominadas fezes moles ou cecotrofos, geradas pela fermentao cecal, o que lhes permite aproveitar mais eficientemente os alimentos com altos teores de parede celular e reingerir nutrientes derivados da atividade bacteriana. BOCA. Como nas outras espcies animais nos coelhos o processo digestivo inicia-se com a apreenso e mastigao dos alimentos (80-120 movimentos por minuto) com a conseqente triturao e insalivao dos mesmos (a diferena do alimento os
cecotrofos so deglutidos ntegros); importante salientar que nos coelhos a saliva apresenta alta atividade enzimtica da -amilase. Depois destes processos digestivos inicias o alimento deglutido passando diretamente atravs do esfago ao estmago. ESTMAGO (1). Mede cerca de 115 mm de comprimento e 75 de largura e apresenta uma capacidade mdia total de 500 ml no coelho adulto, no encontrando-se normalmente vazio; possui dois divertculos bem caracterizados, com um crdia pouco pronunciado e um piloro bastante desenvolvido; (2). Tem uma tnica muscular pouco desenvolvida e pouco contrtil; (3). As secrees estomacais incluem HCl, pepsinognio e mucina. Nos animais lactantes o pH situasse entre 5,5 e 6,0 (tornando-os susceptveis s diarrias) e nos adultos entre 1 e 2. A despeito do baixo pH alguma fermentao ocorre neste local do TGI indicada pela presena de cido lctico decorrente da ao das bactrias nos cecotrofos; INTESTINO DELGADO E RGOS ANEXOS. (1). Comprimento aproximado de 300 cm sendo atingido de maneira total por volta de 9 a 11 semanas de idade; (2). Encontra-se dividido em trs reas funcionais: duodeno, jejuno (maior rea de digesto e absoro) e leo; os processos digestivos que acontecem neste local so similares aos apresentados na maioria das espcies monogstricas. Enquanto que os ruminantes secretam os cidos biliares conjugados com a taurina, nos coelhos estes so secretados com a glicina. Outra peculiaridade que os pigmentos biliares do coelho so principalmente constitudos de biliverdina (como nas aves e anfbios) enquanto que a maioria dos mamferos excretam bilirrubina. INTESTINO GROSSO. (1). Local do TGI bastante volumoso, medindo cerca de 40 cm e com capacidade de aproximadamente 600 ml; est dividido em ceco, clon e reto; (2). A mucosa do ceco bem vascularizada e rica em clulas mucoprodutoras e para a absoro. A poro proximal do ceco se relaciona com a juno leocecoclica, bastante importante na fisiologia deste e do clon. A poro distal do ceco apresenta um apndice vermiforme (de 13 cm de cumprimento em coelhos de 4 meses de idade) que contem numerosas clulas linfides (relacionadas com a secreo de ons bicarbonato, tamponantes dos cidos graxos volteis produzidos durante a fermentao cecal); neste apndice onde ocorre a fagocitose bacteriana. A apendicotomia reduz, significativamente, os nveis de vitamina B12 no contedo cecal; (3). O clon tem um tamanho aproximado de 130 cm e est dividido em quatro regies: clon anterior proximal (5-15 cm), clon posterior proximal (de maior comprimento que o anterior), fusus coli (com comprimento muito pequeno, de 3 a 4 cm, este stio responsvel pela separao das fezes duras e moles) e clon distal (apresenta nmero elevado de clulas mucoprodutoras); (4). Aps a digesto dos nutrientes do alimento no intestino delgado, os resduos ainda no digeridos passam atravs da vlvula ileal e seguem parte ao clon proximal e parte ao ceco. O ceco tem importante papel na digesto do coelho devido fermentao que ali acontece, excreo seletiva de fibra e reingesto do
contedo cecal (cecotrofia). No contedo cecal destacam-se as bactrias anaerbias, especialmente os bacilos no esporulados gram positivos, assim como a falta de lactobacilos, alm do mais neste local no existe uma populao significativa de protozorios provavelmente devido ausncia de substratos adequados como amido e acares solveis. O ceco proximal realiza movimentos anti-peristlticos e contraes no seu incio e fim as quais impulsionam parte deste maneira ao ceco, onde o seu contedo misturado continuamente graas s contraes rpidas que acontecem desde a base do apndice at a juno ileocecoclica e vice-versa. Estes movimentos so responsveis pelo maior fluxo das partculas maiores e com pouco lquido at o clon proximal e do maior fluxo das partculas menores, os microorganismos e os lquidos ao ceco. O material contido no clon perde gua e rapidamente eliminado em resposta a uma estimulao nervosa. No coelho as fezes so classificadas como moles (cecotrofos) e duras. Os cecotrofos so produzidos depois que o contedo cecal foi submetido durante algumas horas ao das bactrias. Sua produo inicia-se em resposta passagem completa do contedo cecal pela vlvula ileal. Por sua vez as fezes duras so constitudas pelas partculas maiores e sua eliminao sempre precede contraes simples e amplas do ceco e clon proximal, com rpida movimentao destas atravs do clon distal e reto; conseqentemente, o coelho capaz de excretar rpida e seletivamente a fibra da dieta, retendo no ceco por tempo prolongado as fraes solveis e as partculas menores dos alimentos. A estratgia de produzir dois tipos de fezes capacita ao coelho a utilizar dietas altas em forragens, com parede celular de baixa digestibilidade e simultaneamente utilizar os demais constituintes das forragens. Nos coelhos a capacidade de digesto da parede celular dos alimentos significativamente inferior aos ruminantes e os herbvoros com fermentao psgstrica como o cavalo. Esta diferena se deve ao rpido trnsito do alimento, ao mecanismo que impede a entrada ao ceco de partculas fibrosas maiores e as limitaes que tm as bactrias cecais por fontes ricas em energia uma vez que ao ceco somente chega o material residual do processo digestivo. Os cidos graxos volteis (AGV) formados no ceco durante a fermentao resultam da atividade microbiana que, por sua vez, depende da capacidade de utilizao dos microorganismos pelos constituintes da parede celular dos alimentos. Embora as propores de AGV no contedo cecal variam por inmeros fatores (tipo de dieta, estratgias de oferecimento do alimento, tempo aps a refeio) se pode dizer que esto entre 60 e 70% para o cido actico, vo de 15 a 20% para o cido butrico e entre 10 e 15% para o propinico. Os AGV produzidos so absorvidos no ceco e clon proximal, porm uma quantidade considervel eliminada juntamente com as fezes cecotrofas o que permite aos coelhos diferirem as fezes duras das moles. Existem sugestes de que os nveis de cido butrico tenha relao com a velocidade de trnsito da digesta: os aumentos na proporo molar deste AGV geram aumento do tempo de reteno do alimento no TGI, diminuio dos movimentos peristlticos e, em conseqncia, transtornos digestivos. Alguns autores tm sugerido, por um outro lado, que determinadas concentraes cecais de AGV poderiam regular a ingesto de
alimentos devido presena de receptores qumicos sensveis ao acetato, lactato e propionato, alm de induzirem a eliminao de cecotrofos. Os microorganismos cecais tambm exibem a capacidade de sntese de vitaminas do complexo B e a vitamina K. O fenmeno de cecotrofia permite ao coelho que sejam cobertas as necessidades destas vitaminas com exceo da piridoxina (B6), cianocobalamina (B12) e tiamina. No contedo cecal dos coelhos possvel achar quantidades significativas de uria oriunda do prprio material procedente do leo, que por sua vez vinha da dieta, e do sangue; contudo, a uria do alimento tem importncia restrita visto que grande parte desta degradada ainda antes de atingir o ceco. Os microorganismos contidos no ceco esto capacitados para transformar a uria em amnia e utiliz-la na sntese de protena microbiana ou para absorv-la atravs das paredes da mucosa devido que a mucosa do ceco apresenta a enzima urease, que hidrolisa a uria em amnia; estes achados sugerem que nas dietas para coelhos possvel substituir at 25% do nitrognio total utilizado para mantena por fontes de nitrognio no protico. Contudo, a sntese de protena a partir da fermentao microbiana significa para o animal adulto cerca de 22% dos gastos totais de energia de mantena. Como resultado da sntese microbiana o contedo cecal de aminocidos totais e essenciais superior ao fornecido pela dieta; grande parte destes aminocidos podem ser reaproveitados pelos coelhos utilizando a cecotrofia; esta particularidade representa para o coelho o fornecimento dirio, adicional ao alimento, de 13,8 g de protena/kg de peso vivo/dia. 4. EQINO. BOCA. (1). Inclui os agentes da apreenso: dentes, lbio superior ( bastante mvel) e lngua; (2). Dentes (a) O movimento da mandbula tanto vertical quanto lateral; (b) A mandbula superior mais ampla do que a inferior, assim, a mastigao s pode acontecer simultaneamente num lado da boca. (3). Saliva (a) Contm pouca enzima -amilase, alm de ter pouco tempo para atuar; a saliva pouco eficiente quanto a digesto enzimtica; conseqentemente, sua funo principal a de umedecer fortemente os alimentos; (b) Sua secreo estimulada pelo atrito (ao mecnica) do alimento sobre a membrana mucosa interna da bochecha; (c) Nos cavalos maduros podem ser secretados at 35 litros de saliva por dia; (d) Trs pares de glndulas esto particularmente desenvolvidas: as submaxilares, as sublinguais e as partidas (estas ltimas s funcionam durante a mastigao); (4). A apreenso dos alimentos auxiliada pelos lbios, dentes e os movimentos da cabea; (5). A ingesto rpida de um alimento celulsico, finamente esmagado, junto a uma mastigao incompleta e a uma fraca produo de saliva, podem provocar graves perturbaes gstricas;
10
(6). A digesto nos eqinos explicada em grande parte do que passa na cavidade bucal. ESFAGO. (1). Tubo comprido (1,25 a 1,50 m) que vai desde a boca at o estmago no lado esquerdo do pescoo; (2). O vu do palato muito desenvolvido e a presena de s um tipo de movimento peristltico fazem que a deglutio seja um processo irreversvel e difcil. O aparecimento de rejeio gstrica pelas narinas rara e, caso acontea, indica a presena de uma dilatao esofgica ou, ento, rompimento do crdia e, portanto, a morte do animal. ESTMAGO. (1). De pouco volume (15 a 20 l) quando comparado com outras espcies, o que obriga refeio de pequenas quantidades de alimento vrias vezes por dia; (2). No apresenta intensa atividade muscular como em outras espcies o que faz, portanto, que o alimento se arranje com freqncia formando camadas; isto pode produzir no eqino grandes problemas digestivos originados no estmago; (3). possvel distinguir duas partes: a esquerda, ou de grande tuberosidade, revestida de um epitlio que contm essencialmente glndulas que secretam muco e a direita que apresenta glndulas gstricas secretoras de HCL e pepsina; (4). A alimentao abundante determina o esvaziamento do estmago de 6 a 8 vezes por dia. Em conseqncia de uma passagem relativamente rpida dos alimentos por este local do TGI e de um pH no muito elevado, nos eqinos e limitada a digesto gstrica. (5). Desde o fim da refeio a parte dos alimentos que fica no estmago, principalmente no caso dos glucdios, sofre uma rpida digesto microbiana. A posterior secreo de HCl acarreta uma queda no pH estomacal, a hidrlise das protenas em molculas menores mediada pela atividade da pepsina e, em conseqncia, a diminuio da atividade microbiana. INTESTINO DELGADO. (1). Tem mais de 20 metros de comprimento e apresenta uma elevada capacidade de armazenagem (60-70 l); semelhante ao do suno, porm no apresenta vescula biliar e, consequentemente, h despejo direto da blis dentro do duodeno; (2). Normalmente o estmago esvazia-se lentamente durante as horas que seguem refeio (3 a 8 horas) produzindo a chegada de pequenas quantidades de contedo estomacal ao duodeno as quais permanecem por pouco tempo neste local; (3). No intestino delgado a secreo biliar e pancretica continua e a intestinal abundante e rica em inmeras enzimas; contudo, a digesto neste local encontrase limitada pelo pouco tempo de permanncia do alimento; INTESTINO GROSSO. (1). Explica a maior parte da capacidade total do TGI (acima de 60%); nesta parte do TGI onde o alimento permanece durante maior tempo; (2). Est dividido em ceco, clon menor ou flutuante, clon maior ou dobrado e reto; (3). Ceco e clon maior:
11
(a). O ceco separado do intestino delgado pela vlvula leo-cecal e do clon dobrado pelo orifcio ceco-ilaco; (b). O clon maior por sua vez dividido em quatro compartimentos: clons ventrais (direito e esquerdo) e clons dorsais (esquerdo e direito); (c). O ceco contm uma flora ativa, constituda por bactrias, semelhante da populao microbiana que existe nos ruminantes e que cumpre com as seguintes funes: A quebra bacteriana da celulose e outros carboidratos para produzir cidos graxos volteis (actico, propinico, butrico) o que faz que o eqino seja tido como animal capaz de utilizar alimentos fibrosos; Sntese bacteriana de vitaminas solveis em gua; Sntese de protena microbiana; (d). No ceco se apresenta absoro de alguns cidos graxos volteis (AGV); visto que as protenas e outras molculas de elevado peso produzidas no ceco e clon maior no esto submetidos ao das secrees digestivas, parece que seja limitada sua utilizao. (4). Clon menor. rea principal de reabsoro dgua do contedo intestinal; (5). Devido que o intestino grosso dilatado com o material ingerido, a impactao pode ocorrer facilmente. 5. SUNO. BOCA rgo inicial do TGI que apresenta trs rgos acessrios:
(1) Lngua. Apreenso, mistura e deglutio; (2) Dentes: Apreenso e mastigao; (3) Glndulas salivares: trs pares (submaxilares, sublinguais, partidas) que secretam
a saliva, sendo que seus componentes so: (a) gua: umedece o alimento consumido e ajuda aos mecanismos comprometidos com o gosto; (b) Mucina: ajuda na lubrificao para a mastigao; (c) Sais de bicarbonato: atuam como tampo para a regulao do pH do estmago; (d) Enzima (amilase salivar). Presente nos sunos, mas duvidosa a sua importncia na digesto dos carboidratos dado que o alimento rapidamente engolido. Deve-se considerar ainda que o pH da saliva 7,3, um pouco acima daquele considerado ideal para a atividade da enzima. (4). Os sunos utilizam o lbio inferior, dentes e lngua na apreenso dos alimentos; (5). Na boca os alimentos permanecem durante pouco tempo, no sendo suficientemente mastigados e insalivados; esta a primeira causa da pouca adaptao dos sunos ao consumo de alimentos ricos em fibra e volumosos. ESFAGO. (1). Tubo muscular oco que transporta o alimento desde a boca at o estmago; o material ingerido mobilizado por uma srie de contraes musculares relacionadas com as ondas peristlticas; (2). Entre o esfago e o estmago existe o esfncter cardial que impede o retorno do alimento desde o estmago para a boca, exceo dos casos de vmito.
12
ESTMAGO. rgo muscular digestivo oco em forma de pra que tem quatro regies: esofgica, cardaca, fndica e pilrica; extremadamente semelhante ao do cavalo. No entanto a primeira regio nos sunos totalmente desprovida de glndulas secretoras, as outras trs possuem glndulas (cardacas, fndicas e pilricas) disseminadas por toda sua camada mucosa que produzem diferentes secrees. (1). Funes: (a) Armazenagem do alimento ingerido. O seu volume normal est entre 7 e 8 litros em um animal adulto; (b) Apresentao de movimentos musculares que produzem o rompimento fsico das partculas do alimento e sua mistura com as secrees gstricas; (c) Secreo de sucos digestivos (cido clordrico, pepsina e renina) com uma composio intermediria entre os carnvoros e os herbvoros, o que pode definir sua posio como sendo um animal de tipo alimentar onvoro; (d) Torna possvel a passagem dos alimentos ao intestino; (2). O pH do estmago aproximadamente 2; (3). O material que deixa o estmago chamado de quimo. INTESTINO DELGADO. (1). Est dividido em trs sees: (a) Primeira seo: Duodeno. Recebe as secrees do pncreas, fgado (a blis armazenada na vescula biliar) e das paredes do intestino, alm de ser o local ativo da digesto. (b) Seo mdia: Jejuno. Ativa para a absoro de nutrientes. (c) Seo posterior: leo. Tambm ativa para a absoro de nutrientes. (2). As paredes do intestino delgado esto arranjadas em uma srie de projees em forma de dedo chamada villi as quais servem para aumentar a rea de absoro. Cada villi possui uma pequena artria e uma veia, junto com um tubo de drenagem do sistema linftico (lacteal). As veias finalmente drenam dentro do sistema portal o qual vai diretamente ao fgado; o sistema linftico se esvazia atravs do conduto torcico dentro de veia cava. (3). O pH do contedo do intestino delgado prximo de 6 a 7. INTESTINO GROSSO. (1). Est dividido em trs sees: (a) Primeira seo: Ceco; seu tamanho vria de maneira considervel entre as diferentes espcies; no suno tem pouca importncia fisiolgica; (b) Seo mdia: Clon; ocupa a maior parte do intestino grosso; (c) ltima seo: Reto. (2). Funes (a) Local de absoro dgua e minerais; (b) Secreo de alguns elementos minerais como o Ca++; (c) Reservatrio para o armazenagem de contedos no digeridos no TGI;
13 (d) Fermentao bacteriana e, portanto, de digesto de material fibroso, sntese de
protena e de algumas vitaminas solveis em gua e vitamina K. IV. PROCESSO DIGESTIVO. Os processos digestivos nas espcies no ruminantes implicam tanto aes mecnicas quanto qumicas e enzimticas: 1. Aes mecnicas. Referem-se apreenso do alimento, mastigao e s contraes da musculatura do TGI.
1.1. Apreenso do alimento.
Refere-se a um ato voluntrio, motivado pela sensao de fome e inibido pela saciedade, que realizam as diferentes espcies animais empregando os lbios, lngua, dentes o bico e permite o passagem do alimento do meio externo boca.
1.2. Mastigao.
Consiste na triturao mecnica do alimento de forma a reduzi-lo a partculas de menor tamanho fsico e assim torn-lo acessvel ao ataque das enzimas digestivas. A mastigao se efetiva mediante movimentos verticais e horizontais da mandbula inferior, de forma tal que os dentes de cada mandbula se pressionam mutuamente realizando a diviso e triturao do alimento. Em princpio a mastigao pode ser considerada como um ato voluntrio, mas pode chegar a fazer-se de maneira mecnica mediante ato reflexo.
2. Aes qumicas e enzimticas.
Os princpios nutritivos dos alimentos consistem de polmeros de unidades simples unidas por enlaces de diversa natureza. O amido, por exemplo, constitudo por cadeias de unidades de glicose ligadas por ligaes glicosdicas. A formao dos nutrientes que necessita o organismo animal (glicose, por exemplo) consistir no rompimento dos princpios nutritivos (amido, por exemplo) em suas unidades mais simples; isto se realiza, fundamentalmente, pela ruptura de suas ligaes, mediante a adio de uma molcula de gua (hidrlise). Esta hidrlise pode ser efetuada graas ao das enzimas ora produzidas pelo TGI ora pelos microorganismos prprios e presentes nele. As funes do TGI esto reguladas pelo sistema nervoso e pelos hormnios gastrointestinais. A inervao extrnseca do TGI est configurada pelas fibras prganglionares do sistema parassimptico, as fibras ps-glanglionares do sistema simptico e as fibras aferentes das vsceras; estas ltimas tm por funo transmitir, atravs dos estmulos gerados nos diferentes locais do trato, os impulsos ao sistema nervoso central para o controle da atividade das vsceras e do prprio comportamento do animal como por exemplo o associado com a fome e a saciedade. A enervao prpria do trato, ou sistema nervoso intrnseco, est sob a ao do sistema nervoso entrico mediante os complexos ganglionares submucoso ou de Meissner e o mientrico ou de Auerbach, os que esto distribudos ao longo das paredes do trato desde a faringe at o nus at a cloaca. O sistema nervoso intrnseco essencial para
14
as funes gastrointestinais, porm este no pode mant-las dentro de um nvel adequado sem a assistncia do sistema nervoso extrnseco. O fornecimento de sangue no TGI varia nas diferentes regies, contudo bem mais rico naqueles segmentos onde existe elevada atividade secretria e de absoro como por exemplo no intestino delgado. As contraes do TGI e as secrees digestivas, tanto qumicas como enzimticas, tm lugar como resposta aos mecanismos nervosos e hormonais no animal. No caso dos mecanismos nervosos o ato reflexo que produz o estmulo secreo pode ser simples ou condicionado. O primeiro se produz quando acontece o contato entre as paredes do TGI e as partculas da digesta ou os produtos qumicos originados pelo processo digestivo; no entanto, os reflexos condicionados so gerados a partir das percepes sensoriais do animal (vista, ouvido, tato, olfato). O mecanismo de controle hormonal se inicia a partir de um ato reflexo; neste caso o hormnio induz, posteriormente, a secreo local ou externa das enzimas ou os compostos qumicos (HCl, bicarbonato de sdio, por exemplo) do TGI. O controle hormonal do TGI est sob a ao dos seguintes grupos de componentes qumicos: 1. Hormnios peptdicos: Gastrina, secretina, colecistoquinina, bombesina, bulbogastrona; 2. Peptdeos hormonais: Polipeptdeo pancretico, peptdeo intestinal vasoativo, peptdeo libertador da gastrina e peptdeo inibidor da gastrina; 3. Neurotransmissores: Acetilcolina, norepinefrina, alguns peptdeos e purinas como o liberador da gastrina. A lista de peptdeos reguladores do TGI grande; dela fazem parte, alm dos j relatados, os seguintes: Substncia P, neurotensina, TRH, metionina-enquefalina, endorfinas, fator libertador da corticotropina, glicentina, polipeptdeo libertador do hormnio do crescimento, peptdeo tirosina-tirosina, neuropeptdeo Y, hormnio libertador da tirotropina e enteroglucagon. O TGI das espcies animais domsticas produz um nmero importante de enzimas geradas a partir de quatro possveis vias: 1. Glndulas anexas ao TGI: Salivais e pncreas, por exemplo; 2. Glndulas encontradas nas prprias paredes em diferentes locais do TGI (abomaso, estmago, pro-ventrculo, papo); 3. Clulas libertadas de maneira permanente mediante os processos de restituio das paredes do TGI; 4. A partir das populaes de microorganismos que conseguem se adaptar s condies cambiantes do TGI. As enzimas hidrolisam os seus substratos descompondo-os em unidades mais simples; as principais enzimas originadas a partir das trs primeiras vias do TGI esto registradas na Tabela 1. Nos monogstricos a flora microbiana do TGI est composta principalmente por bactrias e, possivelmente, protozorios. A flora bacteriana compreende grande quantidade de espcies com atividade enzimtica diferente, sendo as mais importantes:
15
1. Bactrias celulolticas: requerem para crescer os AGV de cadeia ramificada produzidos pelas bactrias proteolticas. Algumas das bactrias celulolticas mais destacveis so: Ruminococcus albus, R. flavefaciens, Bacteroides succinogenes, Butyrivibrio fibrisolvens; 2. Bactrias hemicelulolticas: Butyrivibrio fibrisolvens, Ruminococcus sp, Bacteroides sp, principalmente ruminicola; 3. Bactrias pectinolticas: Butyrivibrio fibrisolvens, Bacteroides ruminicola, B. succinogenes, Peptostretococcus; 4. Bactrias amilolticas e de carboidratos solveis: Streptococcus bovis, Bacteroides amylophilus, B. ruminicola, Succinomonas amylolytica, Selenomonas ruminantium, clostridium butyricum e alguns grupos de Butyrivibrio fibrisolvens e B. succinogenes; 5. Bactrias que utilizam acares: Todos os grupos anteriores de bactrias utilizam os dissacardeos originados a partir da degradao dos polissacardeos. As bactrias que no tm a capacidade de degradar e fermentar os polissacardeos, como por exemplo com os Lactobacillus, dependem dos monossacardeos liberados por outras bactrias. Os principais grupos de bactrias que formam parte deste grupo so: Lactobacillus, Borrelia sp, Succinovibrio dextrinosolvens, E ruminantium; 6. Bactrias que utilizam cidos orgnicos: Selenomonas ruminantium, Veilonella alcalescens, Peptostreptococcus elsdenii, Vibrio succinogenes; 7. Bactrias proteolticas: Bacteroides amylophilus, B. ruminicola, Selenomonas sp, Butyrivibrio sp, Succinovibrio sp, Borrelia sp, Megasphera olsdenii; 8. Bactrias ureolticas: Streptococcus faecium, Succinovibrio dextrinosolvens, Selenomonas sp, Bacteroides ruminicola; 9. Bactrias lipolticas: Veillonella ulcalesceus, Anaerovibrio lipolitica, Butyrivibrio sp, B. fibrisolvens, Treponemas brianti, eubacterium sp, Fusocillus sp, Micrococcus sp; 10. Bactrias metanognicas: Metanobacterium, Methanosarcina barker, Metamicrobium mobile. Em conjunto esta flora possui -amilase, celulase, hemicelulase, pectinase, protease, desaminase, lipase e muitas outras enzimas. Durante a mastigao o alimento alm de ser triturado impregnado pela saliva. A saliva um lquido incolor, espumoso e opalescente que contem 98% de gua e 2% de compostos orgnicos (mucina, enzimas) e sais minerais (carbonatos, ons). A saliva exerce efeito umectante sobre os alimentos secos abrandando-os, dissolve as substncias spidas dos alimentos, contribui com a sensao de gosto, lubrifica o alimento e inicia o processo digestivo enzimtico mediante a amilase salivar (-amilase ou ptialina) que produzida por diversas classes de glndulas salivares que vertem seu contedo ao interior da cavidade bucal.
16
TABELA 1. PRINCIPAIS ENZIMAS DIGESTIVAS NOS NO RUMINANTES. Local de secreo Glndulas salivais Nome Condies para ser ativa on cloreto; pH: 6.6 e 6.8 Substrato Produtos principais de sua atividade Maltose, maltotriose, 1-6 glicosdeo Mono e diglicerdeos; cidos graxos livres Polipeptdeos
amilase (ptialina)
Lipase
Hidrlise dos enlaces -1,4 das ligaes glicosdicas do amido Triglicerdeos
Pepsina
Glndulas do estmago, abomaso e proventrculo
Renina
HCl; pH: 1,5-2,2 Ligaes peptdicas adjacentes ao aminocidos aromticos (fenilalanina, tirosina, triptofano) e dicarboxlicos (cido glutmico e asprtico) Ca; pH: 4 Casena Protena coagulada Meio cido. Por Triglicerdeos isso limitada sua atividade no estmago pH: 7 Hidrlise enlaces -1,4 das ligaes glicosdicas do amido Ligaes ster Sais biliares, primrias dos fosfolipdeos, colipase; pH: 8 triglicerdeos Sais biliares steres de colesterol Fosfolipdeos Enteroquinase Polipeptdeo
Lipase gstrica
amilase (ptilina)
Mono e diglicerdeos; cidos graxos livres Maltose, maltotriose, 1-6 glicosdeo Mono e diglicerdeos; cidos graxos livres Colesterol livre; cidos graxos livres Lisofosfolipdeo s; cidos graxos livres Pequenos
Lipase
Colesteril esterhidrolase Fosfolipase A2 Tripsina
17
Pncreas excrino
Quimotripsina
intestinal; pH: 5.2-6.0; converso autocataltica Tripsina; pH: 8
com um grupo arginina ou lisina terminal Polipeptdeo com aminocido aromtico terminal Hidrlise das ligaes peptdicas dos oligopeptdeos Catalizam a quebra da ligao acarH3PO4 do RNA Catalizam a quebra da ligao acarH3PO4 do DNA Completam a ao das carboxipetidase s hidrolisando os polipeptdeos no extremo do aminocido livre Dipeptdeos Nucleotdeos Nucleosdeos
polipeptdeos; dipeptdeos Polipeptdeos; dipeptdeos Peptdeos; aminocidos livres Nucleotdeos
Carboxipeptidas Tripsina es Ribonucleases
Desoxirribonucl eases Aminopeptidase s
Nucleosdeos
Peptdeos; aminocidos livres
Dipetidases Polinucleotidase s Nucleosidase
Lipase
Triglicerdeos
Intestino delgado
Sacarase Maltase Lactase Isomaltase Fosfatase
pH: 5-7 pH: 5.8-6.2 pH: 5.4-6.0 pH: 6.0-6.5 pH: 8.6
Sacarose Escindem a maltose Lactose Isomaltose Fosfatos
Aminocidos Nucleosdeos; H3PO4 Purinas e pirimidinas; fosfato de pentosas Di e moniglicerdeos; cidos graxos livres; colesterol Frutose; glicose Glicose Glicose; galactose Glicose Fosfato livre
18
A umidade do epitlio da boca se mantm graas secreo continua da saliva, a qual ocorre mesmo que no existam estmulos para a sua secreo; porm, pouca a importncia quantitativa deste tipo de secreo. A presena dos alimentos na boca, a sensao de gosto, a excitao mecnica das mucosas, a mastigao e qualquer outra percepo sensorial que chegue ao hipotlamo e que o animal associe com a ingesto de alimentos, so talvez os atos reflexos que maior influencia tm sobre a produo e secreo da saliva. O bolo alimentcio (mistura de alimento ingerido, mastigado e homogeneizado com a saliva) passa desde a boca ao estmago ou papo nas aves, atravessando a faringe e o esfago; este ltimo se comunica com o estmago atravs de um esfncter chamado crdia, o qual permanece geralmente fechado e s se abre com a chegada da onda peristltica durante a deglutio. Nos alimentos slidos a deglutio ocorre em trs etapas ou fases denominadas: bucal, farngea e esofgica. No entanto a primeira corresponde a um ato voluntrio, as outras duas fases so originadas pelos movimentos peristlticos das fibras musculares estriadas que se propagam em forma de onda (movimentos involuntrios). Quando se tem ingesto de alimentos lquidos ou muito fluidos este so projetados rapidamente at o esfago mediante a contrao dos diversos msculos que existem na regio; o trnsito destes alimentos pelo esfago se deve a este impulso e fora da gravidade. Quando o bolo alimentar chega ao estmago submetido ao das suas secrees; estas consistem da mistura de duas classes distintas de secrees produzidas por dois tipos de glndulas situadas na mucosa gstrica: As glndulas cardacas e pilricas elaboram uma secreo rica em mucina e com pH bsico; por sua vez as glndulas da regio pilrica segregam o hormnio gastrina. A mucina forma uma cobertura na mucosa estomacal e resistente ao resto da secreo gstrica; As glndulas fndicas contm duas classes diferentes de clulas; umas denominadas principais ou peptdicas e as outras chamadas de parietais ou oxnticas; en tanto as primeiras elaboram uma secreo de natureza enzimtica, as parietais segregam o HCl. As secrees gstricas (HCl, pepsina, mucina) so estimuladas e inibidas mediante inmeros mecanismos: Os estmulos que chegam ao hipotlamo atravs dos sentidos (gosto, por exemplo) estimulam as secrees gstricas (fase ceflica da secreo gstrica); Quando o estmago est vazio suas paredes se encontram prximas devido a uma certa tenso de sua camada muscular intermediria. Com a chegada de alimento ao estmago suas paredes se relaxam dispondo-se em camadas concntricas cada vez mais prximas do crdia o que j constitui um estmulo das secrees gstricas (estmulo fsico). A gua e os produtos inicias do processo digestivo encima das protenas (peptdeos e alguns aminocidos) tambm estimulam as secrees gstricas prvio o estmulo da mucosa glandular da regio pilrica para produzir e liberar o hormnio gastrina (estmulo qumico). Este hormnio abandona o estmago, absorvido no intestino delgado e cai na corrente sangnea voltando por esta mesma via para estimular s glndulas estomacais de secreo. Desta forma produzida a chamada fase gstrica das secrees estomacais.
19
A mistura das secrees gstricas com os bolos alimentcios tem lugar graas aos movimentos peristlticos os que se iniciam na regio do crdia e, posteriormente, se propagam numa onda em direo ao antro pilrico onde uma nova onda muito mais enrgica empurra os bolos at o ploro. O ploro permanece fechado enquanto o contedo estomacal no alcana um certo nvel de fluidez e acidez e, portanto, este contedo refluir ao corpo do estmago para continua-se misturando com as secrees gstricas at atingir as condies que permitam a apertura do ploro e, em conseqncia, sua sada, mediante uma onda peristltica, ao intestino delgado. O ploro, por sua vez, de novo se fecha devido distenso do duodeno perante a entrada do contedo estomacal, a queda imediata do pH intestinal e a liberao do hormnio enterogastrona pela mucosa intestinal como resposta presena de gordura neste local do TGI. Este hormnio chega, por via sangnea, mucosa estomacal inibindo seus movimentos e secrees; desta forma produz-se a fase intestinal do processo digestivo no estmago. O HCl desenvolve vrias funes importantes no estmago:
Destroi a maior parte dos microorganismos que chegam com o alimento ao
estmago;
Solubiliza os sais minerais, favorecendo sua absoro; Com a liberao do seu H+ produz-se a queda do pH estomacal o que, por sua vez,
provoca a desnaturao das protenas ingeridas aumentando seu tempo de permanncia neste local do TGI e gera as condies necessrias para ativar os complexos enzimticos. A pepsina secretada inicialmente como zimgeno, ou a forma inativa da enzima, sendo chamada de pepsinognio; desta forma se evita a destruio das glndulas secretoras do estmago; a liberao de um peptdeo inibidor pela ao do HCl fazem ativa esta pr-enzima convertendo-a em pepsina; posteriormente a prpria pepsina ativa as novas molculas inativas produzidas e liberadas no estmago. A pepsina, alm de hidrolisar as ligaes peptdicas adjacentes aos aminocidos aromticos e exercer forte ao sobre os dicarboxlicos, possui ainda uma atividade coaguladora sobre as protenas do leite. Nos mamferos produzida a zimosina ou renina enquanto os animais estejam em lactao; depois desta ser suspensa e visto que mudam as condies estomacais quanto ao pH, se reduz consideravelmente sua produo e diminui sua atividade devido a que ela requer um pH prximo neutralidade. Esta enzima tem grande importncia no que diz respeito da digesto gstrica sobre a protena do leite porquanto precipita a casena formando um coalho, prolongam seu tempo de permanncia no estmago e, portanto asseguram uma digesto enzimtica mais estvel, permitem uma passagem contnua e prolongada dos produtos digestivos gerados at o intestino delgado evitando, desta forma, a sobrecarga excessiva deste local do TGI com protenas sem digerir ou com molculas ainda de elevado peso molecular como so os polipeptdeos. No estmago, fundamentalmente a digesto representada pela hidrlise parcial das protenas do alimento em polipeptdeos a cargo da pepsina ou a renina. Neste local do TGI tambm tem lugar a absoro de H20, sais minerais e alguns dos monossacardeos livres contidos no alimento.
20
A mucosa intestinal ao ser estimulada pela acidez do contedo procedente do estmago secreta pr-secretina, precursor do hormnio secretina, o qual liberado para a corrente sangnea e ao atingir a circulao pancretica estimula as clulas do pncreas a secretarem ons carbonatos, algumas enzimas, mucina e outros compostos orgnicos. As secrees pancreticas ricas em ons e compostos orgnicos junto blis contribuem na neutralizao do pH do contedo intestinal a fim de que este seja alvo das enzimas achadas neste local. As enzimas pancreticas mais importantes so a amilase, lipase, proteases e peptidases. Ao contrrio de todas as enzimas pancreticas, a amilase liberada ao intestino delgado em estado ativo. A lipase pancretica atua conjuntamente com a lipase gstrica, mas sua ao mais importante. As proteases e peptidases so secretadas sob a forma inativa (pr-enzimas): tripsinognio, quimotripsinognio e procarboxipeptidases A e B. Estas formas so ativadas ao liberar-se o polipeptdeo inibidor mediante a ao da enteroquinase, uma enzima especfica produzida na mucosa intestinal. Portanto, a enteroquinase ativa o tripsinognio que se transforma na sua forma ativa a tripsina, quem, por sua vez, ativa o quimotripsinognio e as procarboxipeptidases a quimotripsina e carboxipetidases, respectivamente. A blis produzida pelo fgado e liberada no duodeno, em resposta a estmulos qumicos, hormonais e nervosos, atravs do duto biliar. Com exceo dos equdeos, todas as espcies animais tm uma vescula -biliar-, muito evoluda nos coelhos, que serve como reservatrio das secrees biliares. Na blis encontram-se em dissoluo os seguintes compostos: Pigmentos biliares: principalmente bilirubina e biliverdina. Produzidos no fgado a partir da destruio das porfirinas contidas nas hemcias; Lpides saponificveis: so esterides que emulsionam as gorduras formando gotas minsculas com superfcie acessvel ao ataque enzimtico, sendo os mais destacveis os cidos biliares; Mucina e outras pequenas quantidades de substncias orgnicas. A presena de digesta no duodeno provoca a estimulao mecnica e qumica do intestino gerando mecanismos de resposta que so tanto nervosos quanto hormonais. A mucosa duodenal, por exemplo, libera o hormnio enteroquinina que estimula a secreo do suco entrico. A parede intestinal secreta o suco entrico, que constitudo por sua vez por dois tipos de secrees: Suco duodenal: produzido pelas glndulas de Brunner, localizadas no duodeno. Trata-se de uma secreo alcalina com pH entre 8,2 e 8,9, rica em mucina e amilase, enzima que atua sobre os -polmeros dos monossacardeos ainda no atacados; Suco intestinal: secretado pelas glndulas de Lieberkhun, rico em enzimas digestivas de atividade prxima neutralidade como: a oligoglucosidase, maltase, lactase, sacarase, lipase, aminopeptidases, dipeptidases, nucleotidases e ainda outras enzimas de menor importncia quantitativa (fosfatases, colestinases e mucinases). No intestino delgado, portanto, a digesto se completa pela ao combinada das enzimas fornecidas pelo pncreas e o epitlio do intestino delgado.
21
MECANISMO DE ABSORO DOS NUTRIENTES. O alimento ingerido progride ao longo da boca, faringe e esfago mediante a deglutio. Posteriormente, a mistura do bolo alimentcio com as secrees gstricas vai se deslocando em direo caudal ao longo do intestino delgado. O contedo intestinal prossegue por intermdio de movimentos peristlticos, de segmentao, pendulares e rotatrios como resposta presso deste sobre a parede intestinal o que gera seu avano e mistura com as secrees prprias e que desembocam neste local do TGI. Ao longo do intestino delgado tem lugar a absoro de grande parte dos nutrientes e a energia contidos no alimento. Esta absoro se v facilitada pela grande quantidade de villi que possui a mucosa intestinal as quais esto em permanente contato com o contedo intestinal. Cada villi contem abaixo uma camada de clulas, um vaso linftico e a ramificao de uma arterola que termina em uma pequena vnula. A absoro implica diferentes tipos de mecanismos de transporte: Diretos: Ativo ou contra o gradiente de concentrao; Passivo ou por difuso Indiretos ou pinocitose. A membrana celular possui uma invaginao que vai englobando as molculas at encerr-las no citoplasma formando um vaculo. Este um mecanismo muito freqente nos mamferos nas primeiras horas de vida aps o nascimento que permite a absoro de molculas complexas e de elevado peso mesmo sem serem ainda submetidas aos processos de digesto, como acontece, por exemplo, com as -globulinas do colostro.
As ondas peristlticas do intestino delgado se detm ao chegar ao final do leo. Quando o contedo intestinal chega vlvula leo-cecal parte dele passa ao ceco durante o tempo em que permanea aberto seu esfncter; assim que este se feche interrompida a passagem do contedo e o leo volta a sua posio original. Devido que esse processo se repete vrias vezes a passagem do contedo intestinal ao ceco em forma descontnua. No ceco o contedo intestinal submetido a movimentos peristlticos e antiperistlticos; embora estes sejam mais lentos, quando comparados com os do intestino delgado, so suficientes para provocar sua homogeneizao e avano at o clon. Durante o trnsito pelo ceco e clon, os resduos do alimento e os constituintes endgenos produzidos ao longo do TGI que ainda no foram digeridos so postos em contato com a flora microbiana ali presente e com a mucina secretada pelo intestino grosso. O tempo de permanncia do contedo intestinal no ceco varivel dependendo da espcie animal e o tipo de parede celular presente nos alimentos ingeridos: nas aves e sunos esta pode ser praticamente inexpressiva caso que a fibra do alimento for constituda por componentes parietais muito indigerveis; no caso dos herbvoros a digesta permanece por mais tempo neste local do TGI. No ceco e clon se mistura a digesta, procedente do intestino delgado, com a mucina produzida pelas glndulas e as clulas da mucosa, ajudando a formar assim um contedo intestinal mais compacto. Neste local do TGI continua a ao das enzimas
22
secretadas anteriormente pelo intestino delgado; contudo, o processo digestivo neste local caracterizado pela fermentao microbiana sobre os resduos do alimento resistentes s enzimas, as clulas descamadas ao longo do todo o TGI e sobre os prprios microorganismos mortos. O produto mais importante da ao digestiva dos microorganismos so os cidos graxos volteis (AGV) que so absorvidos diretamente ali por simples difuso. No ceco e clon tambm so absorvidos amnia, alguns aminocidos e vitaminas sintetizadas pelos prprios microorganismos (K e algumas do complexo B). Posteriormente as fezes so transportadas ao reto mediante movimentos peristlticos; a dilatao deste local no TGI estimula a gerao de movimentos musculares voluntrios que abrem os dois esfncteres que o comunicam com o nus provocando sua expulso. Com a defecao pode se dizer que conclui o processo digestivo para um grande nmero das espcies animais, incluindo o homem; a exceo mais marcante est com os animais que praticam a coprofagia j que com a ingesto de seus prprios excrementos repete-se o processo digestivo. As fezes expulsas ao exterior esto compostas dos seguintes constituintes: Partculas de alimento ingerido que resistiram s diferentes aes do processo digestivo ou que no foram absorvidas; Resduos das secrees produzidas pelo prprio TGI; Clulas e restos de clulas procedentes dos processos de troca do epitlio do TGI; Microorganismos e resduos da flora prpria do TGI. V. REFERNCIAS BIBLIOGRFICAS DE APOIO SOBRE A DIGESTO COMPARADA EM NO-RUMINANTES. 1. Bergman, E.N. Contributions of VFA from the gastrointestinal tract in various species. Physiological Reviews. 70 (2): 567-590. 1990. 2. Corring, T.; Juste, C.; Lhoste, E.F. Nutritional regulation of pancreatic and biliary secretions. Nutrition Research Review. 2: 161-180. 1980. 3. Cranwell, P.D. Development of the neonatal gut and enzyme systems. p 99-154. In: Varley, M.A. The neonatal pig. Development and survival. CAB International. 1995. 4. Hintz, H.F.; Cymbaluk, N.F. Nutrition of horse. Annual Review of Nutrition. 14:243-268. 1994. 5. Holst, J.J. The neuro-endocrine control of digestive processes. In: Digestive Physiolgy in the Pig. Proceeding of the 3rd International Seminar. Copenhagen 16th-18th may 1985. National Institute of Animal Science. Editors: Just, A., Jorgensen, H., Fernandez, J.A. p. 17-34. 6. Johnson, L.R. Regulation of gastrointestinal mucosal growth. Physiological Reviews. 68 (2): 456-502. 1988.
23
7. Low, A.G. Nutritional regulation of gastric secretion, digestion and emptying. Nutrition Research Review. 3: 229-252. 1990. 8. Macari, M., Furlan, R.L., Nakaghi, L.O. Anatomia e histologia funcional do trato digestivo. In: Curso de Fisiologia da Digesto e Absoro de Aves. Junho 7-8, 1992. Santos, So Paulo. Brasil. Fundao Apinco de Cincia e Tecnologia Avcolas. P. 1-17. 9. Marounek, M. et al. Distribution of activity of hidrolytic enzymes in the digestive tract of rabbits. Britishy Journal of Nutrition. 73 (3): 463-469. 1995. 10. Maxwell, F.J.; Stewart, C.S. Microbiology of the gut and the role of probiotics. p. 155-186. In: Varley, M.A. The neonatal pig. Development and survival. CAB International. 1995. 11. Mei, N. Intestinal chemosensitivity. Physiological Reviews. 65 (2): 211-237. 1985. 12. Moog. F. El revestimiento del intestino delgado. Investigacin y Ciencia. Enero 1982. p. 92-102. 13. Moran, E, T. Jr. Comparative nutrition of fowl & suine. The gastrointestinal systems. Canada. 1982. 253 p. 14. Nabuurs, M.J.A. Microbiological, structural and functional changes of the small intestine of pigs at weaning. Pigs News nad Information. 16 (3): 93N-97N. 1995. 15. Nitsan, Z. et al. Growth and development of the digestive organs and some enzymes in broiler chicks after hatching. British Poultry Science. 32: 515-523. 1991. 16. Pinchasov, Y. Early transition of the digestive system to exogenous nutrition in domestic post-hatch birds. British Journal of Nutrition. 73 (3): 471-478. 1995. 17. Pluske, J.R. et al. Nutrition of the neonatal pig. p. 187-238. In: Varley, M.A. The neonatal pig. Development and survival. CAB International. 1995. 18. Yamauchi, K.E.; Isshiki, Y. Scanning electron microscopic observations on the intestinal villi in growing white leghorn and broiler chickens from 1 to 30 days of the age. British Poultry Science. 32: 67-78. 1991.
GUA. INTRODUO. Tradicionalmente, nutriente tem sido definido como o constituinte ou grupos de constituintes de um alimento que tem a mesma composio qumica geral e ajuda na
24
manuteno da vida. Atualmente nossa interpretao sobre o significado de nutriente vai alm desta definio inicial visto que nela ns tambm incluirmos substncias que no so necessariamente originadas no alimento (vitaminas produzidas sinteticamente, sais inorgnicos preparados quimicamente e aminocidos sintetizados biogeneticamente). No podemos ter a certeza de que a lista dos nutrientes conhecida at hoje encontrados nos alimentos ou nos tecidos animais tenha todos os nutrientes necessrios para o funcionamento do organismo animal. Embora o estudo dos nutrientes menos complexos como a gua quanto a sua descrio, funes fisiolgicas especficas e seus requerimentos bastante simples, , tambm junto ao oxignio, o mais esquecido. Por sua vez, no que tem a ver com os nutrientes que servem ao fornecimento de energia (a qual por sua vez no pode ser classificada como um nutriente propriamente dito) ou que desempenham inmeras funes nos processos metablicos de liberao e uso de energia, mais complexa a compreenso dos processos nutricionais. Estas razes explicam porque geralmente mais fcil realizar a caracterizao dos alimentos a partir de sua composio qumica que a classificao dos nutrientes que o compem. O grfico 1 resume a forma clssica como representada a composio qumica dos alimentos. GUA. A.GERAL. 1. Junto com o oxignio a gua o constituinte mais abundante e importante para a manuteno da vida:
a. Representa de 65 a 85% do peso corporal no animal ao nascimento e entre 45 e
60% do mesmo na idade madura e explica entre 90 e 95% do contedo total do sangue. O teor de gua corporal varia pouco dentro de cada espcie, mas depende da idade do animal, das variaes no peso corporal e, consequentemente, de seu contedo de gordura. Os registros das tabelas 1 e 2 mostram que a porcentagem da gua corporal diminui significativamente com a idade do animal e com o aumento da deposio de gordura corporal; estes mesmos registros sugerem que no existem variaes significativas no que diz respeito do contedo de gua no plasma sangneo diante as mudanas na idade do animal.
25
Protena Compostos nitrogenados Fontes de nitrognio no protico Matria Orgnica Alimento Matria Seca Carboidratos Extrativos no nitrogenados Fibra bruta Vitaminas Hidrosolveis Macromineral Matria Inorgnica (Minerais) Micromineral Lpides
Aminocidos no essenciais Aminocidos semiessenciais Aminocidos essenciais
Simples Compostos Vitaminas Monossacarides Oligossacarides Polissacarides Polissacarides Essenciais Essenciais Provavelmente essenciais Alguns podem ser txicos Outros no parece que sejam essenciais
gua
GRFICO 1. COMPOSIO QUMICA DOS ALIMENTOS.
26
TABELA 1. Contedo de gua corporal e no plasma sangneo de aves Leghorn Branca de acordo com a idade dos animais. Idade dos animais (semanas) 1 2 3 4 6 8 16 32 Contedo de gua (valores expressos em %) Corporal 95.5 96.3 96.1 95.8 95.8 95.5 95.1 94.6 Plasma sangneo 85.2 68.7 67.1 68.9 59.2 65.9 48.7 55.0
TABELA 2. Variaes nos contedos de gua e gordura corporal de acordo com o peso vivo de sunos tipo carne. Peso vivo dos animais (kg) 15 20 40 60 80 100 120 Porcentagem da carcaa gua 70.4 69.6 65.7 61.8 58.0 54.2 50.4 Gordura 9.5 10.1 14.1 18.5 23.2 27.9 32.7
b. A gua est distribuda de forma heterognea no corpo animal de modo a manter o equilbrio dinmico entre os comportamentos do organismo: (a). gua intracelular representa mais de 50% do peso vivo e constitui o meio onde ocorrem as reaes biolgicas; (b). gua extracelular achada principalmente nos fludos intersticiais, plasma sangneo, linfa e fludos sinovial e crebro-espinhal constitui aproximadamente 20% do peso corporal; (c). gua presente na urina e trato gastrointestinal. Estando o plasma em constante equilbrio com o fludo intersticial, a composio de ambos permanece inalterada em todas as situaes. c. O corpo dos animais domsticos e do homem pode perder praticamente toda a gordura e mais da metade da protena e sobreviver, enquanto a perda de 10% da gua pode resultar em morte. A capacidade de suportar a privao de gua e de perd-la varia segundo as espcies: as poedeiras Leghorn, por exemplo, podem sofrer a restrio de 10 a 20% no consumo de gua sem afetar o seu desempenho; no entanto, se a restrio exceder 20% ocorrero graves prejuzos
27
na sua produo. J os jumentos podem suportar a perda de 30% de sua gua corporal sem graves riscos para sua vida; B. PROPRIEDADES DA GUA. 1. Alta constante dieltrica; 2. Baixa viscosidade: esta propriedade permite a passagem de gua e das substncias nela dispersas at os mais finos vasos capilares do organismo sem elevadas exigncias para o organismo; 3. Boa tenso superficial: propriedade da gua que permite que ela se mantenha fortemente ligada superfcie de qualquer outra substncia; 4. Alto calor especfico o que permite a absoro de uma enorme quantidade de calor proveniente do trabalho muscular; 5. Alta condutibilidade trmica: permite o transporte do calor absorvido para a superfcie do corpo ou para o lume do intestino. C. FUNES PRINCIPAIS DA GUA. 1. Constituinte dos fludos corporais: Crebro-espinhal: protege o sistema nervoso; Sinovial: lubrifica as juntas; Auricular: transporte de ondas sonoras; Intra-ocular: importante no processo da viso; Amnitico: protege o feto. 2. Regulao da presso osmtica intracelular realizada atravs da ingesto e eliminao de gua e/ou eletrlitos na urina; 3. Regulao da temperatura corporal devido a seu alto calor especfico e a uso dos mecanismos de evaporao cutnea e pulmonar. O alto calor especfico apresentado pela gua significa dizer que cada grama de gua necessita de muito calor para elevar muito pouco sua temperatura ou que ela capaz de absorver grandes quantidades de calor com uma elevao mnima da temperatura corporal; 4. Principal componente das secrees animais: leite, ovos, secrees hormonais e enzimas digestivas, fetais e do crescimento; 5. Participa nos processos de hidrlise na digesto, a absoro dos nutrientes no trato digestivo, o transporte de todos os componentes qumicos do organismo, o metabolismo intermedirio nos tecidos e a excreo de seus produtos o que a faz ser o solvente universal; 6. Componente corporal com maior taxa de reciclagem; 7. Dispersante ideal devido ao seu poder ionizante; 8. Boa condutora da eletricidade o que importante na transmisso neural; 9. Participante da homeostase orgnica mantendo o equilbrio cido-base. D. FONTES DE GUA PARA O ANIMAL. A cobertura das necessidades de gua proveniente de trs origens:
1. gua para consumo direto ou gua de bebida.
28
a principal fonte para os animais, devendo apresentar certas caractersticas que afetam sua qualidade e, consequentemente, seu consumo direto.
a. Fatores que afetam o consumo de gua de bebida:
(1). Temperatura e umidade relativa do ambiente: normalmente o consumo de gua aumenta quando o animal est por fora das suas condies de conforto. O aumento do calor ambiente leva a um incremento da transpirao que, por sua vez, eleva as necessidades de gua consumida; sob condies de baixa temperatura tambm acontece aumento quanto s exigncias de gua de bebida porquanto ocorre o catabolismo das protenas orgnicas. Os efeitos das variaes na temperatura ambiente sobre o consumo de gua tambm esto associados com a condio fisiolgica do animal: frangos de corte, por exemplo, dobram o consumo de gua quando a temperatura ambiente passa de 22 para 32o C entanto que as poedeiras triplicam o consumo de gua quando a temperatura passa de 21 para 37o C; (2). A prpria temperatura da gua: os animais diminuem a ingesto voluntria de gua quando a temperatura desta menor que 6o C ou superior a 36o C; (3). Fatores da dieta: (a) Consumo da matria seca: dentro dos intervalos de temperatura de conforto o consumo de gua est diretamente relacionado com o consumo de matria seca; (b). Altos teores de gua no alimento reduz o consumo desta; (c). Altos teores de sais e protena aumenta o consumo de gua. (4). Tipo de sistema urinrio: as aves exigem menor quantidade de gua do que os mamferos em percentagem de peso vivo devido ao tipo de excreo do nitrognio urinrio. Entanto as aves excretam cido rico, que necessita menor quantidade de gua para sua eliminao, os mamferos precisam de mais de 100 g de gua para eliminar to s 1 g de uria; (5). Idade e estado fisiolgico do animal: para cada kg de peso vivo os animais jovens necessitam mais gua que animais de maior idade. Por sua vez, as vacas em lactao precisam mais gua (em mdia para cada litro de leite produzido, so necessrios 873 g de gua). As porcas em lactao tambm consomem mais gua que as gestantes e estas mais que os animais em crescimento. As aves poedeiras, sob condies normais, fora do perodo de postura consumem 166 ml de gua/dia; este valor aumenta a 306 ml visto que a produo de um ovo exige de 37 g de gua; (6). A qualidade dgua afeta o seu prprio consumo: (a). gua de boa qualidade para consumo deve ser incolor, inspida e inodora, com pH entre 7,0 e 7,2 (nveis de pH acima de 7,2 indicam alcalinidade o que sugere a necessidade de se pesquisar os nveis de clcio e magnsio) e livre de contaminao bacteriana. (b). gua de boa qualidade deve ter menos do que 2500 mg/litro de slidos dissolvidos; (c). As guas que tenham acima de 1g de sulfatos/litro podem produzir diarrias; (d). A presena de elementos como nitratos, flor, ferro e molibdnio em excesso so extremadamente txicos. Nveis de 100 a 200 ppm de nitratos na gua, por exemplo, so potencialmente txicos.
29
b. Consumo aproximado de gua em animais maduros e no estressados:
Aves: Duas partes de gua para cada parte de matria seca consumida; Ovinos: 3-10 litros; Sunos: 5-10 litros; Bovinos e eqinos: 35-50 litros.
2. gua contida nos alimentos. a. Varia conforme o tipo de alimento (estdio fisiolgico, processamento,
armazenagem, etc.); (1). Nos gros pode variar desde valores abaixo de 8% at valores acima de 30%; (2). Nas forragens varia desde valores abaixo de 5% nos fenos secos at valores acima de 90% no caso dos capins novos. (3). A quantidade de gua dos alimentos pode influenciar a quantidade de gua livremente ingerida pelo animal e o valor energtico do alimento. Neste ltimo caso pode dizer-se que com o aumento no percentagem de gua do alimento diminui seu contedo de energia digestvel (ED): assim, por exemplo, o gro de milho, com 12% de gua, tem 3841 Kcal de ED/kg, a mandioca, que pode ter 65% de gua, apresenta 1050 Kcal de ED/kg). b. A gua de chuva sobre os alimentos diminui o consumo direto dgua. 3. gua metablica.
a. Produzida pela oxidao das substncias que contm hidrognio em sua
frmula (carboidratos, protenas e gorduras) nos tecidos animais. As inmeras estimativas realizadas sugerem que a oxidao de 1,4 g de protenas, 1,7 g de carboidratos ou 0,9 de gorduras produz 1 g de gua metablica. 60, 42 e acima de 100 g de gua metablica. A maior quantidade de gua metablica produzida pela oxidao das gorduras est relacionada com o maior contedo de hidrognio em relao ao oxignio dentro da molcula. Por sua vez a quantidade de gua metablica produzida est em relao inversa complexidade do carboidrato: a oxidao da sacarose, por exemplo, fornece 57,9% de gua metablica entanto que a do amido gera 55%. b. Responde somente por 5 a 10% das necessidades dirias de gua dos animais domsticos. Contudo, ela pode suprir as necessidades dirias de alguns animais em hibernao ou em condies desrticas. E. EFEITOS DAS DEFICINCIAS OU RESTRIES DE GUA. 1. Reduz o consumo de alimento e a produtividade; 2. Perdas de peso devido desidratao; 3. Aumenta a excreo de nitrognio e eletrlitos como Na+ e K+. F. PERDAS DE GUA. As perdas de gua tm quatro funes importantes nos animais: 1. Eliminao dos produtos finais do metabolismo, principalmente atravs da urina;
30
2. Regulao da presso osmtica do sangue; 3. Atua na termoregulao pela evaporao cutnea e pulmonar; 4. gua o principal componente das secrees e produtos animais. As perdas de gua podem acontecer atravs de cinco vias: 1. Rins; 2. Fezes: variam em funo da espcie animal, as caractersticas do trato gastrointestinal e o tipo de alimento consumido; 3. Pulmes: o principal fator associado com a regulao da temperatura corporal. A eliminao pulmonar precisa de quantidades suficientes de gua para saturar o ar alveolar, que, por sua vez, depende da umidade do ar inspirado, da temperatura e da ventilao pulmonar. O ar nos pulmes varia em contedo de gua e em temperatura. O ar expirado est prximo da temperatura do corpo e bastante saturado de gua o que representa uma perda de energia considervel para um animal em clima frio e um meio de liberao de calor para um animal sob condies de clima quente visto que 1 g de gua evaporada representa a perda de 576 cal. 4. Via cutnea: a perspirao resulta da difuso de gua atravs do tegumento cutneo entanto que a transpirao a secreo pelas glndulas sudorparas. Com a transpirao no s ocorre a perda de gua mas tambm a de substncias tais como: cloreto de sdio, sulfatos, fosfatos, compostos de enxofre, cido lctico e vitaminas hidrossolveis. 5. Produtos secretados: leite, ovos. G. NECESSIDADES DE GUA. O requisito mnimo de gua de qualquer animal representa a soma das perdas de gua pelo corpo (urina, fezes, evaporao, respirao), mais as perdas associadas reproduo (leite, ovos, pario), mais uma parcela destinada ao crescimento do animal quando jovem que apresenta maior atividade dos tecidos e menor teor de gordura corporal. No existem regras gerais para determinar as necessidades de gua. Geralmente para realizar uma estimativa destas devem ser considerados a superfcie corporal e o metabolismo basal, alm do regime alimentar, sistema de criao, temperatura e umidade do ambiente, exerccio realizado pelo animal, a produo animal e os prprios fatores da qualidade dgua. Na tabela 3 so apresentadas algumas sugestes da quantidade de gua a ser consumida pelos principais animais domsticos. H. REFERNCIAS BIBLIOGRFICAS DE APOIO. ANDRIGUETO, J.M. Nutrio animal. So Paulo. Nobel. 1982. 395p. ANDRIGUETO, J.M. Nutrio animal. 3a ed. So Paulo. Nobel. 1986. 425p. BERTECHINI, A.G. Nutrio de monogstricos. ESAL/FAEPE. Lavras. Minas Gerais. 142p. KOLB, E. Fisiologia veterinria. Zaragoza. Acribia. 2a ed. V.2. 1979. NUNES, I.J. Nutrio animal. Escola de Veterinria. UFMG. Belo Horizonte. Minas Gerais. Apostila. 1972. PEIXOTO, R.R & MAIER, J.C. Nutrio e alimentao animal. 2a ed. U Pelotas. Pelotas. 1993. 169p.
31
TABELA 3. CONSUMO DE GUA SUGERIDO PARA DIFERENTES ESPCIES ANIMAIS. CATEGORIA Frangos de corte Frangas Frangas Poedeira Vacas em lactao Vacas e novilhas ao final da gestao Vacas secas e novilhas em gestao Novilhas em idade de inseminao Fmeas desmamadas at inseminao Bezerros lactantes (em baia) Bezerros lactantes (a pasto) Leiteiros Bezerros at 6 semanas Bezerros de 100 ou mais kg de peso vivo At 100C 10-150C 15-200C 20-270C >270C De corte AVES At 8 semanas At 16 semanas 16-22 semanas 90% postura BOVINOS CONSUMO 1,6-1,8 l/kg rao 2,4 l/kg rao 166 ml/dia 306 ml/dia 62,5 l/animal/dia 50,9 l/animal/dia 45,0 l/animal/dia 48,8 l/animal/dia 29,8 l/animal/dia 1,0 l/animal/dia 11,2 l/animal/dia 40-65 l/dia 6,5 kg/kg MS da rao 3,5 kg/kg MS da rao 3,6 kg/kg MS da rao 4,0-4,2 kg/kg MS da rao 4,5-4,8 kg/kg MS da rao 5,6 kg/kg MS da rao 8-9 l/100 kg P.V. 45 l/animal/dia Benedetti (1987) AUTOR NRC (1984)
Andrigueto (1982)
CO
32
Animais jovens Fmeas em lactao CAPRINOS
2-3 l/kg MS da rao 4 l/kg MS da rao 3,5 l/kg de leite produzido 1,43 kg/kg leite produzido 145,6 g/kg 0,75 COBAIA Com forneciment o de forragem verde e fresca Sem a suplementa o de forragem verde COELHO NRC (1981) Recomenda o francesa Recomenda o francesa NRC (1972)
50-100 ml/animal/dia
250-1000 ml/animal/dia
Animal adulto Fmeas antes do parto Fmeas em lactao EQINOS Cavalos em descanso Cavalos em trabalho pesado SUNOS Adultos Leites de 5-8 semanas Sunos em terminao
0,25 l/animal/dia 1 l/animal/dia Segundo o nmero de lparos 37 l/animal/dia 57 l/animal/dia 1,9-2,5 kg/kg de rao seca 20 l/100 kg P.V. 7 l/100 kg P.V./dia
Andrigueto (1986)
Crowell (1985)
NRC (1988)
DIGESTO DOS CARBOIDRATOS NOS NO-RUMINANTES. INTRODUO
33
O nome carboidrato derivado do francs hidrate de carbone e faz referncia aos componentes que na natureza contem carbono, hidrognio e oxignio, sendo que o hidrognio e o oxignio sempre aparecem na mesma proporo como encontrada na gua (2:1). Embora definio seja til para caraterizar este grupo de compostos qumicos, ela ainda limitada porquanto o fsforo, enxofre ou nitrognio tambm podem fazer parte da estrutura dos carboidratos. Os carboidratos so feitos de compostos aldedicos ou cetnicos com mltiplas hidroxilas e, portanto, so definidos como poliidroxialdedos ou poliidroxicetonas, com frmula geral (CH2O)n, sendo que n 3. Embora muitos dos carboidratos comuns apresentam na frmula a proporo 1:2:1 entre os tomos de carbono, hidrognio e oxignio, alguns no cumprem com esta regra. I. FUNES. Os carboidratos constituem a maior parte da matria orgnica na terra devido a suas mltiplas funes em todas as formas de vida. � Primeiro, os carboidratos servem de reservas energticas, alimentos energticos e intermedirios metablicos: � amido nas plantas e o glicognio nos animais so carboidratos que podem ser rapidamente mobilizados para gerar glicose (principal fonte alimentar para a produo de energia; � ATP e muitas coenzimas so derivados glicdicos fosforilados. Nas plantas, os carboidratos so originados a partir do gs carbnico (CO2) atmosfrico e dgua atravs de uma das mais importantes reaes qumicas que existem natureza, a fotossntese. Embora a reao subentende a formao de grande nmero de produtos intermedirios, pode ser, simplesmente, representada da seguinte maneira: 6CO2 + 6H2O + 673Kcal C6H12O6 + 6O2 glicose Os carboidratos dos vegetais so, por sua vez, utilizados pelo animal como fontes de energia para os seus processos orgnicos e, assim, toda a vida animal depende tambm da fotossntese. Nutricionalmente falando, o major problema que se apresenta aos animais no est associado com a disponibilidade de carboidratos na natureza e sim com sua capacidade para digeri-los e absorv-los. Ao se comparar com as protenas e gorduras, os carboidratos fornecem menos energia para o metabolismo. Contudo, esta uma concluso provisria. Os carboidratos acabam sendo os nutrientes que mais contribuem com o fornecimento de energia na alimentao animal visto que so encontrados em maior quantidade nas plantas (70 a 75%), so altamente digestveis e sua participao normalmente alta nas dietas.
34
� Segundo, os carboidratos ribose e desoxirribose formam parte do esqueleto estrutural do DNA e do RNA. A flexibilidade na conformao desses anis de oses importante no armazenamento e na expresso da informao gentica. � Terceiro, alguns tipos de carboidratos so elementos estruturais das paredes celulares de plantas, bactrias e dos exoesqueletos de artrpodes. De fato, a celulose, o principal constituinte das paredes celulares das plantas, o composto orgnico mais abundante da biosfera. � Finalmente, os carboidratos so ligados a muitas protenas e lipdeos. � As unidades glicdicas da glicoforina do s hemcias um revestimento aninico altamente polar; � Unidades glicdicas nas superfcies das clulas participam de maneira importante no reconhecimento de clula a clula durante o desenvolvimento. II. ESTRUTURA. Os carboidratos esto constitudos (em % por peso molecular) de C (40%), H (7%) e O (53%), sendo que os tomos de C esto arranjados em cadeias s quais esto ligados o O e H e estes dois, por sua vez, tm semelhana qumica com a molcula de gua. As molculas de carboidratos mais simples podem se apresentar sob duas configuraes absolutas: D e L, dependendo da posio que toma o grupamento hidroxila (OH) do penltimo carbono. Importante lembrar que, em uma projeo de Fisher de uma molcula, os tomos unidos a um tomo de carbono assimtrico por ligaes horizontais esto na frente da pgina, e os unidos por ligaes verticais esto atrs (Fig. 1).
H | C=O | HCOH | HCOH | H D-gliceraldedo
H | C=O | OHCH | HCOH | H L-gliceraldedo
Figura 1. Representao das formas D e L do gliceraldedo.
35
Para os monossacardeos com mais de um tomo de carbono assimtrico, os smbolos D e L referem-se configurao absoluta do carbono assimtrico mais distante do grupamento aldedo ou cetona. Nos animais somente os carboidratos de configurao D so metabolizados; no entanto, suas formas L so menos freqentes na natureza sendo metabolizados principalmente pelo microorganismos. Em soluo as molculas de carboidratos podem girar a luz polarizada para a direita (dextrgeras) ou para a esquerda (levgiras), isto representado pelos sinais (+) e (-); assim, por exemplo, a D (+) dextrorrotatria e a L (-) levorrotatria. Por causa desta propriedade, algumas vezes os carboidratos so chamadas de dextrose e levulose. III. CLASSIFICAO DOS CARBOIDRATOS. A Tabela 1 traz um resumo da classificao dos carboidratos mais significativos em nutrio animal. De uma maneira simples os carboidratos podem ser classificados em acares e no acares. � Acares: so carboidratos relativamente simples, de baixo peso molecular e solveis em gua. Neste grupo encontram-se os monossacardeos e os oligossacardeos. TABELA 1. Classificao esquemtica dos carboidratos mais significativos em nutrio animal. Trioses Gliceraldedo C3H6O3 Diidroxiacetona Eritrose Tetroses Eritrulose C4H8O4 Ribose Pentoses Ribulose C5H10O5 Xilose Monossacardeos Xilulose Arabinose Glicose Hexoses (Dextrose) C6H12O6 Frutose (Levulose) Galactose Manose ACARES Heptoses Sedoeptulose C7H14O7 Glicose-Frutose Sacarose Galactose-Glicose Lactose Dissacardeos Glicose-Glicose Maltose Oligossacardeos Trealose Glicose-Glicose Celobiose Trissacardeos Rafinose Tetrassacardeos Estaquiose
36
Pentosanas Homopolissacarde os Hexosanas
Arabanas (Arabinanas) Xilanas Glicanas Frutanas Mananas Galacturanas Glicosaminas
Amido, glicognio, celulose, dextrinas Inulina, levana cido pctico quitina
NO ACARES
Hemiceluloses Gomas Mucilagens Heteroploissacarde Substncias os pcticas Sulfopolissacardes Aminopolissacarde s
cido hialurnico, condroitina, heparina Os aucares simples que na sua estrutura possuem um grupo aldedo (-CHO-) so denominados de aldoses e os que possuem o grupo cetona (-CO-), so conhecidos como cetoses. Nutricionalmente, a aldose mais importante a D-glicose entanto que a cetose mais importante a D-frutose. Em razo destes grupos os monossacardeos podem ser oxidados, reduzidos, ou substitudos; fornecendo derivados de importncia metablica e nutricional. � No acares: constitudo por carboidratos mais complexos, de alto peso molecular, insolveis em gua ou que formam solues coloidais. Deste grupo fazem parte os homopolissacardeos e os heteropolissacardeos. Uma outra forma, mais complexa, de classificar os carboidratos baseia-se no nmero de tomos de carbono por molcula e no nmero de molculas por composto. 1. CLASSIFICAO DOS CARBOIDRATOS QUANTO AO NMERO TOMOS DE CARBONO. � � � � � Trioses (C3 H6 O3 ) Tetroses (C4 H8 O4 ) Pentoses (C5 H10 O5 ) Hexoses (C6 H12 O6 ) Heptoses (C7 H14 O7 ) 2. CLASSIFICAO DOS CARBOIDRATOS QUANTO AO SEU NMERO DE MOLCULAS. 2.1. MONOSSACARDEOS. Os monossacardeos so slidos cristalinos, incolores, alguns apresentam sabor doce, muito solveis em gua e insolveis em solventes no polares (clorofrmio, benzeno e ter).
37
Embora os monossacardeos ou acares simples sejam os glicdeos mais simples (esto constitudos por uma unidade de carboidrato), so importantes por sua alta capacidade de fornecer esqueletos de carbono para a sntese de outros compostos como, por exemplo, os aminocidos no essenciais; por sua grande afinidade pelo cido fosfrico e, consequentemente, a formao de compostos de alta energia (ADP, ATP); por sua participao na constituio do DNA e RNA e por sua presena em certas plantas como glicosdeos txicos. O esqueleto dos monossacardeos est constitudo por uma cadeia no ramificada de tomos de carbono unidos entre si por ligaes simples. Um dos tomos de carbono unido a um tomo de oxignio por dupla ligao formando um grupo carbonila e cada um dos demais tomos de carbono est ligado a um grupo hidroxila. Caso o grupo carbonila esteja numa extremidade da cadeia, o monossacardeo um aldedo sendo chamado de aldose, mas se este grupo estiver em qualquer outra posio o monossacardeo se caracteriza como uma cetona e chamado de cetose. A presena destes grupamentos ativos, aldedo e cetona, d aos monossacardeos a propriedade de serem redutores ou no, e isto faz com que possam ser oxidados, reduzidos ou substitudos, fornecendo derivados de importncia metablica e nutricional. Exceto a diidroxicetona, todos os monossacardeos comuns que existem na natureza contm um ou mais carbonos assimtricos ou quirais, o que os converte em ismeros ticamente ativos. Os estereoismeros dos monossacardeos podem ser todos relacionados a um composto de referncia, o gliceraldedo, que tem uma forma D e L. Entretanto, como muitas das aldoses tm dois ou mais centros quirais, os prefixos-D e L so usados em referncia configurao do carbono quiral mais distante do tomo de carbono da carbonila. Quando o grupo hidroxila desse tomo de carbono mais distante projeta-se para a direita na frmula, ela designa um D-acar. De maneira similar podemos escrever as estruturas de todos as D-Cetoses at seis tomos de carbono, elas possuem a mesma configurao ao redor do carbono assimtrico mais distante do grupo carbonila. As cetoses so designadas sistematicamente pela insero das letras D no nome da aldose correspondente; por exemplo, D-ribulose a cetopentose correspondente a aldopentose D-ribose. Entretanto, algumas cetoses tem nomes triviais, como a frutose. Em regra, somente os carboidratos de configurao D so metabolizados pelos animais. Alm disso, as forma L so menos comuns na natureza. As formas predominantes da glicose e frutose em soluo no so cadeias abertas; as formas em cadeia aberta destes monossacardeos ciclizam-se formando anis. Em geral, nestes o grupo carbonila no est livre, mas formando uma ligao covalente com um dos grupos hidroxila existente ao longo da cadeia; dependendo da posio que a hidroxila do primeiro toma, recebe a identificao alfa () ou beta (), e quando ocorre a polimerizao de suas molculas, os compostos resultantes apresentam ligaes alfa ou beta. Na D-glicose o aldedo em C-1 na forma em cadeia aberta reage com o grupamento hidroxila em C-5, formando um hemiacetal chamado de piranose devido sua semelhana com o anel hexagonal do pirano; neste caso as formas em anel da Dglicose sero a D-glicopiranose e D-glicopiranosepiranose. De forma semelhante,
38
o grupamento cetnico em C-2 na forma de cadeia aberta da D-frutose pode reagir com a hidroxila em C-5, formando um hemicetal chamado de furanose pela sua semelhana com o anel pentagonal do furano; tambm, neste caso, a forma em anel da D-frutose ser a D-frutofuranose. As formas isomricas dos monossacardeos que diferem entre si apenas na configurao ao redor do tomo de carbono pertencente ao hemiacetal e hemicetal so chamadas formas anomricas ou anmeros e o carbono do hemiacetal e hemicetal ou carbono da carbonila chamado de carbono anomrico (1). 2.1.1. Trioses. So os glicdeos mais simples. So aldedos ou cetonas que tm duas ou mais hidroxilas. O gliceraldedo, uma aldose (contm um grupamento aldedico), e a dihidroxiacetona, uma cetose (contm um grupamento cetnico), so os principais carboidratos deste grupo porquanto so importantes intermedirios na via glicoltica. Embora so pequenas suas concentraes nos fluidos celular e extracelular dos animais, sua taxa de reciclagem extremamente rpida. O gliceraldedo tem um s carbono assimtrico; portanto, h dois ismeros dessa aldose com trs carbonos: o D-gliceraldedo e o L-gliceraldedo (Fig. 1). 2.1.2. Tetroses e heptulose. So carboidratos raros nos tecidos animais. A sedo-heptulose-7-fosfato, entretanto, ocorre como intermedirio na via das pentoses. 2.1.3. Pentoses (frmula geral C5 H10 O5). Os membros mais importantes deste grupo so: � Aldoses (L-arabinose, D-xilose e D-ribose): estes membros raramente ocorrem de forma livre na natureza, exceto como subprodutos da fermentao; � Cetoses (D-xilulose e D-ribulose): so produtos derivados da via das pentoses do metabolismo intermedirio. 2.1.3.1. L-arabinose. Ocorre nas pentosanas como as arabanas. componente das hemiceluloses, goma arbica e outras gomas de exsudao vegetal; tambm encontrada em silagens como resultado da hidrlise. 2.1.3.2. D-xilose. Ocorre nas pentosanas como xilanas. Forma a principal cadeia das hemiceluloses de gramneas.
(1)
Mais um centro assimtrico criado quando a glicose se cicliza. O carbono 1, da carbonila na forma de cadeia aberta, torna-se um centro de assimetria na forma em anel. Este tomo de carbono C-1 chamado de carbono anmero e do mesmo modo as formas e so anmeras. 2.1.4. Hexoses. o grupo mais abundante na natureza. Deste, somente a D-glicose (aldose) e a Dfrutose (cetose) encontram-se como monossacardeos livres sendo que outras hexoses
39
podem ser encontradas como unidades de dissacardeos ou polissacardeos. Ambas, a D-glicose quanto a D-frutose, pertencem srie D porque sua configurao absoluta no C-5 a mesma que a do D-gliceraldedo. 2.1.4.1. D-glicose. Ocorre livre em plantas, frutas, mel, sangue, linfa e liquido cefalorraquidiano. particularmente importante por ser o principal produto final da digesto dos carboidratos nos no ruminantes, fonte imediata de energia para todos os seres vivos e por ser a molcula bsica para a sntese do amido e da celulose. 2.1.4.2. D-frutose. Est de forma livre em folhas verdes, frutos e mel, na sacarose (dissacardeo) e em frutosanas; essencial no metabolismo como frutose-1 e frutose 6-fosfato; geralmente mais lentamente metabolizada que a glicose. Visto que levorrotatria nomeada de levulose. 2.1.4.3. D-manose. No se encontra de forma livre na natureza, mas existe na forma polimerizada, como mananas, o que explica sua presena em fungos, bactrias e leveduras. constituinte comum de glicoprotenas e outros polissacardeos os quais esto presentes no leite e em vrias secrees das mucosas. Este carboidrato no encontrado em quantidade significativa como forma livre em tecidos animais ou fluidos corporais. 2.1.4.4. D-galactose. No est de maneira livre na natureza, exceto como produto de fermentao. Combinada com a glicose forma a lactose, ou acar do leite. Tambm componente de pigmentos antocinicos, galactolipdeos, gomas e mucilagens. Ambas, a D-manose e a D-galactose, so dextrorrotatrias. 2.1.5. Outros monossacardeos.
2.1.5.1. Derivados de monossacardeos.
� steres de cido fosfrico. Exercem papel importante numa ampla gama de reaes metablicas em todos os organismos vivos. Os derivados mais comuns so os formados pela fosforizao da glicose: glicose-1 fosfato e glicose-6 fosfato. � Aminoacares. Em alguns carboidratos no carbono 2 o grupamento -NH2 substitui a oxidrila (OH) originando aminoacares, sendo os mais importantes: a D-glocosamina (est na quitina da concha dos invertebrados e na mucina da saliva e do suco gstrico), a Dgalactosamina (apresenta-se juntamente com o cido glucurnico em sulfato de condoitrina que, combinado em uma glicoprotena, origina um dos principais componentes da cartilagem). � Desoxiacares. So produzidos pela substituio de um grupamento -OH por -H no carbono 2 da estrutura do carboidrato. Deste grupo o desoxiacar mais conhecido a desoxirribose que componente importante do DNA.
40
� cidos acares. As aldoses (L-arabinose, D-xilose e D-ribose) podem ser oxidadas fornecendo cidos. Os mais importantes so os cidos aldnicos (carboxilados no carbono 1), aldricos (olicarboxlicos) e urnicos (carboxilados no ltimo carbono da estrutura). No caso da glicose os derivados mais importantes so os cidos glucnico, glucrico e glucurnico. � lcoois de acares. Aldoses e cetoses podem ser reduzidos produzindo polilcoois. Assim, por exemplo, a glicose produz sorbitol, a galactose dulcitol e a manose (aldose) e frutose (cetose) produzem manitol. � Glicosdeos. So gerados quando o grupo -OH do carbono 1 da glicose substitudo, por esterilizao ou por condensao, por um lcool ou fenol. Embora o termo glicosdeo usado para designar coletivamente tais derivados, os monossacardeos e oligossacardeos so susceptveis mesma reao. Quimicamente o oligo, poli e heterossacardeo so glicosdeos. De um modo geral o termo est associado com glicosdeo txico ou, pelo menos, com glicosdeos que por hidrlise fornecem um resduo que no carboidrato, que, s vezes, chamado de heterosdeos. Dentre dos glicosdeos sobressaem os cianognicos, os quais liberam cianeto de hidrognio (HCN) na hidrlise e que por causa de sua presena em muitas plantas, limitam o seu uso como alimento. Os exemplos destes compostos relatados mais freqentementemente na literatura so: � Linamarina: presente nas sementes de linho (linhaa), feijo-de-java e mandioca; � Vicianina: encontrada na ervilhaca; � Amigdalina: presente nas amndoas amargas e sementes (parte comestvel) do pssego, cereja, ameixa, mas e frutos rosceos; � Durrina: est na parte area do sorgo; � Lotaustralina: contida no trevo branco. 2.2. OLIGOSSACARDEOS. So carboidratos constitudos por 2 a 10 unidades de monossacardeos unidos por ligaes glicosdicas(2), que usualmente so do tipo 1,4, sendo que nas ramificaes, quando presentes, as ligaes so do tipo 1,6. Os carboidratos podem ligar-se uma outra formando di- e polissacardeos. Grande nmero de oligossacardeos tem sido descrito na literatura especializada, sendo que os tecidos animais contm poucos teores deles quando comparados com as plantas. Dentro dos oligossacardeos mais importantes temos: 2.2.1 Dissacardeos (sacarose, maltose, lactose, celobiose e trealose). Formados pela combinao de duas molculas de hexoses com a perda de uma molcula de gua. Trs dissacardeos muito abundantes so a sacarose, a lactose e a maltose. 2.2.1.1. Sacarose.
41
Formada pela combinao dos carbonos anmeros(3) de uma glicose e de uma frutose. Est na cana-de-acar e na beterraba. No um acar redutor. Quando aquecida 160oC produz maltose e aproximadamente 200oC, produz caramelo. 2.2.1.2. Maltose. Formada por molculas de glicose. um acar redutor e no to doce quanto a sacarose. Nos animais um produto da digesto ou a hidrlise do amido ou do glicognio. O produto resultante aps sua secagem a malte, que, por sua vez, usado na fabricao de cerveja, whisky e outras bebidas alcolicas e no alcolicas.
Quando se aquece glicose em metanol anidro contendo HCL, seu tomo de carbono anmero reage com a hidroxila de lcool formando dois acetais: -metil-glicosdeo e -metil-glicosdeo. A nova ligao entre o C-1 da glicose e o tomo de oxignio do metano chamada de uma ligao glicosdica-especificamente, uma ligao glicosdica. As ligaes glicosdicas so facilmente hidrolisadas por agentes cidos, mas so muito resistentes ao de agentes bsicos. (3) O carbono anmero de um carboidrato pode ser ligado ao tomo de N de uma amina por uma ligao N-glicosdica. Este tipo de ligaes em virtualmente todas as biomolculas de ocorrncia natural tm a configurao . A importncia biolgica desse tipo de ligao glicosdica evidente em biomolculas como os nucleotdeos RNA e DNA. 2.2.1.3. Lactose. Formada por ligaes galactose-glicose. o acar do leite, menos doce que a sacarose e facilmente fermentvel, principalmente, por Streptococcus lactis. um acar redutor (4). 2.2.1.4. Celobiose. Trata-se de um carboidrato redutor derivado da hidrlise da celulose que no fermentvel. 2.2.1.5. Trealose. Acar no redutor presente em fungos e algas marinhas. 2.2.2. Trissacardeos. Carboidratos constitudos por trs molculas de hexoses com perda de duas molculas de gua. 2.2.2.1. Rafinose. Como a sacarose tambm est distribuda na natureza, porm em quantidades mais limitadas: est presente nas sementes de algodo (0,8%) e no acar de beterraba, se acumula no melao durante a fabricao do acar. No acar redutor. Sua hidrlise fornece glicose, frutose e galactose. 2.2.3. Tetrassacardeos. Trata-se de carboidratos formados por quatro molculas de hexoses com perda de trs de gua.
(2)
42
2.2.3.1. Estaquiose. Presente nas leguminosas. No um acar redutor. Na hidrlise fornece 2 molculas de galactose, 1 de glicose e 1 de frutose. 2.2.4. POLISSACARDEOS. So polmeros formados por grande nmero de acares simples (mais de 10 unidades). Esto representados por carboidratos complexos que possuem funes de reserva (amido, glicognio) ou de formao de estruturas (celulose, hemiceluloses, pectina, dextrina) e se diferenam entre si pelos monossacardeos que os compem. Pela hidrlise cida completa ou pela ao de enzimas especficas liberam monossacardeos ou seus derivados.
(4). Mono e oligossacardeos contendo um grupamento aldedico ou cetnico livre reduzem indicadores, como, por exemplo, complexos dos ons cprico (Cu2+) forma cuprosa (Cu+). O agente nessas reaes a forma em cadeia aberta da aldose ou cetose. Existem duas espcies de polissacardeos, os homopolissacardeos, que contm apenas um tipo de unidade monomrica, e os heteropolissacardeos, que contm dois ou mais tipos de unidades monomricas. Os carboidratos mais abundantes ingeridos pelos humanos e os animais so os polissacardeos amido e celulose, fornecidos pelos alimentos vegetais, e o glicognio, fornecido pelos alimentos de origem animal. 2.2.4.1. Amido. um homopolissacardeo de reserva que ocorre no interior das clulas vegetais na forma de grandes agregados ou grnulos. O amido especialmente abundante em razes como a batata, e em algumas sementes, como nos gros de cereais, mas a capacidade de sintetiz-lo est presente na maioria das clulas vegetais. As molculas de amido so altamente hidratadas devido ao fato de possurem muitos grupos hidroxila expostos; por esta razo o amido quando extrado dos grnulos com gua quente forma solues coloidais turvas compostas por dois tipos de fraes: � Amilose: a parte mais solvel. Compe de 10 a 20 % do amido. um polmero formado por cadeias longas e lineares de D-glicose com ligaes -1�4; � Amilopectina: parte mais insolvel. Compe de 80 a 90 % do amido. um polmero de cadeias lineares de D-glicose com ligaes -1�4 e com pontos de ramificao constitudos por ligaes so -1�6. 2.2.4.2. Glicognio. As clulas animais armazenam a glicose como glicognio que uma forma qumica prontamente mobilizvel de glicose. O glicognio um homopolssacardeo bem grande e ramificado formado por resduos de D-glicose que se repetem inmeras vezes; entanto a parte linear da estrutura est formada por unidades de glicose unidas por ligaes glicosdicas do tipo 1�4, as ramificaes so constitudas tambm por ligaes glicosdicas porm do tipo 1�6, ocorrendo uma vez a cada 10 unidades. A presena destas ramificaes na estrutura do carboidrato serve para aumentar sua solubilidade, e tornar suas unidades de glicoses mais facilmente mobilizveis.
43
O glicognio armazenado aumenta a quantidade de glicose disponvel para o animal entre as refeies ou durante a atividade muscular. Os dois locais principais de armazenamento de glicose so o fgado, onde pode chegar a representar 7% da massa mida do rgo, e o msculo esqueltico. A sntese e a degradao do glicognio so importantes porque regulam o nvel de glicose no sangue e fornecem uma reserva de glicose para a atividade muscular energtica. 2.2.5. CARACTERSTICAS ESTRUTURAIS E QUMICAS DOS COMPONENTES DA PAREDE CELULAR VEGETAL. Devido importncia que tem os constituintes da parede celular vegetal (polissacardeos estruturais e outros componentes qumicos) na alimentao animal se faz necessrio introduzir uma outra breve reviso sobre este assunto. 2.2.5.1. Estrutura da parede celular vegetal. Quimicamente a parede da clula vegetal composta de: � Polissacardeos estruturais (Fibrosos): Os polissacardeos estruturais que consistem a parede celular dos vegetais so: � Polmeros das pentoses (arabinose e xilose) e hexoses (glicose, frutose e galactose). Estas unidades bsicas se combinam, dando origem a dois grupos estruturais principais -glicanos e heteroglicanos; � Polissacardeos matriciais. No primeiro grupo se encontra a celulose e no segundo, as pectinas, as hemiceluloses e outros polissacardeos. Ambos os grupos formam a frao insolvel da fibra ou de carboidratos insolveis, tambm chamados de polissacardeos no amilceos. Estes carboidratos, junto com a lignina e certos polissacardeos de reserva, como as gomas constituem a fibra, ou a parte do alimento que no pode ser digerida pelas enzimas digestivas, porm, susceptvel a uma degradao de intensidade varivel, pela atuao microbiana situada em distintas partes do trato digestivo. A fibra da dieta tem sido definida como a parte dos alimentos vegetais que no digerida pelas secrees do trato gastrointestinal. � Substncias de incrustao: � cido ftico � amilides � cutina � glicoprotenas � lignina � silca � taninos Fisicamente, a parede celular vegetal est formada por microfibrillas de celulose, de natureza cristalina, e por uma matriz macromolecular de substncias pcticas, hemiceluloses e lignina. Os diferentes polmeros esto estritamente ligados em uma
44
rede, cuja coeso assegurada por foras intermoleculares e ligaes dbeis (Van der Waals, pontes de hidrognio e ligaes inicas covalentes). A parede celular vegetal se encontra organizada em trs zonas: � Lamela mdia: forma um cimento contguo s clulas do tecido da planta, constituda essencialmente por pectinas; � Parede primria: durante o crescimento da planta a estrutura primeiramente formada que engloba o protoplasma. Se apresenta como uma massa amorfa constituda por hemiceluloses e substncias pcticas em que se dispersam microfibrilas de celulose; � Parede secundria: formada depois ter acabado o crescimento da superfcie da clula. Se apresenta com uma ultraestrutura mais resistente constituda por celulose e hemiceluloses. A principal diferena na parede secundria a grande reduo no contedo de gua, aumento da celulose e presena de substncias como a lignina. Quando iniciada a fase de formao da parede secundria se apresentam os primeiros indcios de acumulao de lignina nos cantos da clula ou onde incide a parede primria; o processo avana rapidamente dos extremos para o meio. As pesquisas tm assinalado que quando a lignificao era completada, a clula morria. A parede primria menos diferenciada do que a parede secundria. A diferenciao determinada pela sua composio em celulose, hemiceluloses e lignina. Com a idade da planta aumenta a concentrao de lignina o que, por sua vez, produz queda na sua digestibilidade. Como a deposio de lignina no ocorre uniformemente, alguns nutrientes da parede ou do contedo celular permanecem susceptveis a digesto ou a fermentao. 2.2.5.1.1. Celulose. Quantitativamente a celulose o componente mais importante da parede celular e como tal a molcula mais abundante da natureza. um homopolissacardeo linear, no ramificado, de alto peso molecular, formado de 10.000 ou mais unidades de D-glicose unidas por ligaes glicosidicas do tipo (14) que fazem s cadeias de D-glicose assumirem uma configurao alongada unidas por fortes ligaes intermoleculares e intramoleculares de pontes de hidrognio formando microfibrilas insolveis. Esta rede fibrilar cristalina impregnada com uma matriz de propriedades cimentantes que consiste de polissacardeos de tipos diferentes e de uma substncia polimrica chamada lignina. A regularidade na estrutura da celulose favorece a formao de uma rede cristalina muito resistente resistente ao dos principais reagentes qumicos, apresentando-se insolvel em meios bsicos, mas pode ser dissolvida por cido sulfrico a 72%, cido clordrico a 40%, cido fosfrico a 84% e cobre amoniacal. 2.2.5.1.2. Hemiceluloses. Representam aproximadamente 40% do material da parede celular dos vegetais constituindo o segundo grupo de carboidratos mais abundante na natureza. Em mdia entre 2 e 12% da matria seca dos vegetais e razes e de 10 a 25% da matria seca das forrageiras e de muitos subprodutos industriais como a polpa de ctricos e de beterraba, so constitudos por hemiceluloses.
45
As hemiceluloses so heteropolissacardeos de estrutura complexa heterognea, composta de grande nmero de polmeros, sendo os xilanos (cadeias lineares de Dxilose) o principal componente, existindo, porm, freqentemente glucomananos (cadeias lineares de D-glicose e D-manose, que podem conter tambm ramificaes de galactose). Um fator comum nas hemiceluloses so as ligaes glicosdicas (14) e a capacidade de serem solubilizadas em cido, alcali. Em plantas forrageiras as hemiceluloses so encontradas freqentemente nas paredes lignificadas e so geralmente insolveis tornando-se solveis quando esta parede delignificada. Segundo a escala de Butler a estrutura de qualquer polissacardeo pode ser definida e classificada segundo as seguintes propriedades: tipos de monossacardeos presentes, tipo de anel formado, posies das ligaes glicosdicas, tamanho e ramificao das cadeias e configurao das ligaes glicosdicas. As hemiceluloses podem ser classificadas em: � Pentosanas (polmeros compostos predominantemente por resduos de pentoses). � Xilanos. So formados de cadeias, lineares de D-xilose unidos principalmente por ligaes (14). Este o principal polmero das hemiceluloses. Os resduos de xilose apresentam-se como D-xilopiranose. Associado a cadeia principal encontramos cadeias laterais curtas de arabinose, cido glucurnico e cido 4-0, metil glucurnico, D-galactose e possivelmente D-glicose. Os xilanos podem ser portanto subdivididos em trs grupos principais baseados no carboidrato presente em suas cadeias laterais: arabinoxilanos, glucoranoxilanos e glucoarabinoxilanos.
�
Arabinogalactanos. Esta pentosana formada por resduos de galactose unidos por ligaes (13) e (16). A arabinose aparece nas cadeias laterais.
� Hexanas (polmeros compostos por resduos de hexoses) � Mananas So encontradas como componentes da parede celular de vrias plantas. As glicomananas so basicamente molculas lineares compostas por resduos de glicopiranose e manopiranose unidos por ligaes (14). � -glucanos So molculas constitudas de resduos de D-glicose unidos por ligaes (1-3) e (14). Distingui-se da celulose por ser solvel em lcalis. Parece ser uma molcula restrita a tecidos imaturos de gramneas e parede celular do endosperma de cereais. Aproximadamente 70% da molcula constituda de resduos de glicose unidos por ligaes (14) e o restante por ligaes (13). � Xiloglucanos
46
So abundantes na parede celular primria de dicotiledneas , mas tm sido tambm isolados em certos cereais. Todos os xiloglucanos so compostos de uma cadeia principal formada de unidades de glicopiranose ligada atravs de ligaes (14). A esta cadeia principal esto ligadas unidades de xiloporanose atravs de ligaes (16). As xiloglucanas de dicotiledneas tm composio bastante varivel contendo acares adicionais como galactose e fucose. As hemiceluloses freqentemente esto associados com a lignina ou seus precursores no apenas em contato como tambm atravs de ligaes covalentes. Estas ligaes podem ser do tipo estr ou ter. A composio das hemiceluloses varia de planta para planta e entre as suas diferentes partes. 2.2.5.1.3. Pectinas. As pectinas, ou substncias pcticas, so polmeros do cido 1,4 -D-galacturnico, que se encontram primordialmente na lamela mdia e parede primria da clula vegetal, participando como elemento cimentante da membrana. A cadeia de cidos galacturnicos se apresenta em forma helicoidal e est associada lateralmente com arabanos e galactanos, estando os grupamentos cidos combinados com sais de clcio e com metil-steres. As pectinas podem ser extradas com uma soluo neutra composta por agentes quelantes (oxalato de amnia, cido etileno-amino-tetrcetico) ou hidrolisada por solues lcalis ou cidas diludas. A presena de pectinas em leguminosas (5-10%) mais marcante do que em gramneas (mdia de 2%), no entanto, est freqentemente acima de 20% nas polpas de beterraba e de ctricos e em alguns frutos como a ma e os ctricos. As pectinas conferem s fibras vegetais uma elevada capacidade higroscpica, determinando em geral um aumento de volume e peso das fezes, bem como de seu grau de viscosidade o que est em estreita relao com o trnsito da digesta. As pectinas ainda se relacionam com a capacidade de troca catinica da fibra vegetal, ligando-se em sua superfcie ions metlicos bivalentes como o clcio, magnsio, zinco e ferro, podendo interferir em sua absoro pr-cecal. A relao com o metabolismo dos lipdios se d pelo processo de adsorso de cidos biliares na matriz da digesta com pctina ao nvel duodenal, indisponibilizando a sua reabsoro ileal reduzindo o pool entero-heptico e, com isto, induzindo a mobilizao do colesterol endgeno para atender a sntese dos cidos biliares. 2.2.5.1.4. Lignina. A lignina parece ser um heteropolmero amorfo condensado de distintos lcoois fenilpropanides cujos precursores so -cumaril, coniferil e o sinapil e os cidos fenlicos e -cumrico, os quais se interligam atravs de ligaes do tipo ter ou covalentes carbono-carbono entre o ncleo benznico e o radical propano ou entre os ncleos benznicos formando uma estrutura tridimensional de elevado peso molecular. A proporo destes componentes irregular entre as espcies vegetais.
47
A lignina biossintetisada nas plantas vasculares por uma seqncia de reaes ramificadoras, que comea com a formao de carboidratos, que so normalmente derivados de CO2 assimilado na fotossntese. CO2 Carboidratos Aminocidos fenilpropanides Derivados do cido cinmico Derivados do lcool cinamil lignina. A polimerizao oxidativa dos monmeros fenilpropanides de caracterstica ao acaso, pelo menos no que observado in vitro. Os produtos de polimerizao tm uma estrutura condensada contendo primariamente ligaes do tipo ter e carbono-carbono entre os fenilpropanides em uma estrutura tridimensional, o que explica porque a lignina to resistente hidrlise. As ligaes da lignina com a celulose so do tipo ter, envolvendo o ncleo benznico da lignina e a hexose da celulose. Esse mesmo tipo de ligao ocorre entre a lignina e a xilose das hemiceluloses; contudo, os demais radicais das hemiceluloses reagem com a lignina atravs de ligaes do tipo ster, razo pela qual a ligao entre a lignina e as hemicelulose mais facilmente rompida por tratamentos qumicos e fsicos do que a existente entre esta e a celulose. A lignina um material depositado em maior proporo durante o espessamento secundrio da membrana celular vegetal; de natureza hidrofbica, insolvel, considerado como material de enchimento, que substitui a gua na parede celular e est incrustada s microfibrilas e os polissacardeos da matriz. Parece ser que sua principal funo est associada cimentao dos polissacardeos componentes da parede celular oferecendo um suporte estrutural planta e proporcionando maior resistncia mecnica contra os microorganismos. Na maior parte dos alimentos concentrados e forragens jovens a lignina est presente em quantidades razoavelmente baixas (menos de 5%). Entretanto, seu contedo aumenta em funo do estado de maturao das plantas e da temperatura ambiente em que se desenvolvem, podendo chegar a conter at 12% em algumas plantas herbceas. Importante assinalar que alguns subprodutos utilizados na alimentao animal, particularmente nos que se incluem os talos, cascas e palhas, contem altas concentraes de lignina. A extrao da frao lignina verdadeira da parede celular apresenta alguns inconvenientes: embora uma parte da lignina seja solvel com as hemiceluloses, somente pode ser extrada pela ao de oxidantes fortes (hipoclorito, permanganato de potssio, trietilenoglicol em meio cido ou parcialmente pelo bissulfito). O que se conhece como lignina bruta representa aquelas substncias diferentes aos carboidratos da parede celular que so insolveis em H2SO4 12M, incluindo, alm da lignina, a cutina, os complexos taninos-protenas e os produtos da reao de Maillard. 2.2.5.1.5. Componentes minoritrios da parede celular. Na parede celular esto presentes outros compostos que podem ou no estar associados aos polissacardeos estruturais e a lignina. Estes constituintes, ainda que em quantidades muito pequenas, podem ter efeitos significativos sobre o comportamento digestivo tanto dos demais componentes da parede celular quanto o contedo celular. Estes componentes so a slica, cutina, taninos e compostos de baixo
48
peso molecular. Os leos essenciais, que fazem parte desse ltimo grupo, esto representados por diversas substncias orgnicas que tm a propriedade comum de solubizar-se em solventes orgnicos. Algumas dessas substncias, que so steres ou teres pertencentes ao grupo das substncias fenlicas e provavelmente ao dos terpenos, possuem especial interesse por exibir atividade antimicrobiana e so txicos para no ruminantes. As substncias terpenides de maior importncia nutricional para no ruminantes so as saponinas e os esterides. Vrios pesquisadores relatam que na parede celular existem dois tipos de fraes qumicas: uma, nomeada fibra solvel, e outra, conhecida como insolvel. Entanto a primeira est constituda pelos polissacardeos no amilceos solveis em gua como o -(1�3) glucano da cevada, as pectinas das frutas, o arabinoxilano do arroz, as galactomanas das leguminosas e os polissacardeos das algas, a fibra insolvel em gua est composta por celulose, hemiceluloses e lignina. IV. DIGESTO DOS CARBOIDRATOS. 1. DIGESTO SALIVAR E GSTRICA. De um modo geral nos no ruminantes a digesto do amido inicia-se na boca mediante a ao da -amilase salivar (ptilaina). Na boca esta enzima apresenta sua atividade mxima diante a presena do on cloreto e pH entre 6,6 e de 6,8. No esfago e na regio esofgica do estmago a ao desta enzima continua durante um tempo e s diminui assim que o alimento comea a se misturar com o suco gstrico, sendo finalmente inativa quando o pH do estmago diminui abaixo de 3,6. Nas aves visto que a insalivao muito escassa por possurem glndulas salivares pouco desenvolvidas, os gros de cereais so embebidos no inglvio antes de serem submetidos digesto enzimtica no pr-ventrculo e moela. Este processo considerado uma macerao fisiolgica, j que d s aves uma maior capacidade de digerir os gros do que os demais no ruminantes. Na regio esofgica do estmago de coelhos, eqinos e sunos e no papo das aves existe uma flora microbiana que digere parte dos carboidratos dos alimentos, gerando, como produtos da fermentao os cidos graxos volteis (AGV) actico, propinico e butrico. 2. DIGESTO NO INTESTINO DELGADO. A medida que saem do estmago ao intestino delgado os produtos parciais da digesto gstrica sobre os carboidratos, so estimuladas as secrees pancreticas, a blis e as secrees das glndulas de Brnner o que produz o aumento progressivo do pH longo do duodeno; quando este atinge um valor em torno de 6 a 7, cria-se o meio adequado para que sejam ativas a amilase pancretica e as carboidrases da mucosa duodenal (sacarase, maltase, lactase.e isomaltase). Nos no ruminantes tanto a amilase salivar quanto a pancretica so enzimas alfaamilases e, portanto, produzem a ruptura por hidrlise dos enlaces tipo 1�4 das ligaes glicosdicas da amilopectina. A ao de ambas estas enzimas geram como produto final a D-glicose, pequenas quantidades de maltose e um ncleo ramificado resistente hidrlise chamado dextrina-limite.
49
A dextrina-limite no pode ser hidrolisada pela -amilase devido que as ligaes (1�6) dos pontos de ramificao no so sensveis ao ataque desta enzima. Para isto necessria a presena de uma enzima de desramificao, a (1�6) glicosidase, que tem a capacidade de hidrolisar este tipo de ligaes e, desta maneira, expor ao da -amilase novos pedaos da cadeia com ligaes (1�4). As carboidrases localizam-se na superfcie externa das clulas epiteliais que revestem o intestino delgado, atuam sobre a sacarose, maltose, lactose e isomaltose e liberam, como produto da sua atividade, os monossacardeos correspondentes. Em conjunto, estes so absorvidos pela membrana das clulas epiteliais da mucosa duodenal, passando posteriormente ao sangue e ao fgado, onde so utilizados nas distintas rotas metablicas para a obteno de energia para suprir os gastos dos trabalhos biolgicos, formao de gordura, etc. Importante salientar que as clulas do intestino delgado tm muitas microvilosidades, caracterstica que produz o aumento da rea superficial do intestino para digesto e absoro de nutrientes. 3. DIGESTO NO INTESTINO GROSSO (CECO E CLON). Tanto os constituintes da dieta no digeridos at o leo terminal quanto os produtos resultantes da digesto que no foram absorvidos nos diferentes locais do intestino delgado, passam ao intestino grosso onde so fermentados pelas bactrias para obter a energia necessria para o seu crescimento e proliferao. A fermentao dos componentes da fibra, assim como a frao amido resistente, podem dar lugar a produo de CO2, hidrognio, metano e cidos graxos de cadeia curta, que no caso dos no-ruminantes apresentam as seguintes propores: TABELA 2. Proporo molar dos cidos graxos volteis produzidos pelos noruminates. PROPORO MOLAR DOS CIDOS GRAXOS VOLTEIS ESPCIE ACTICO (%) PROPINICO (%) BUTRICO (%) SUNOS 60 - 77 17 - 21 5-7 COELHOS 60 - 70 10 - 15 15 - 20 EQINOS 70 - 75 18 - 23 5-7 Desde o ponto de vista quantitativo o processo de fermentao no intestino grosso faz com que o amido e os acares sejam digeridos em sua totalidade e que os carboidratos estruturais o sejam em maior ou menor proporo dependendo de seu nvel e, sobretudo, do tipo de carboidrato estrutural presente nas dietas. A produo de cidos graxos volteis a partir da fermentao dos resduos que escaparam do processo digestivo no estmago, duodeno, jejuno e leo, que chegam ao intestino grosso, pode constituir em funo da espcie animal e seu estado fisiolgico, em uma importante contribuio ao metabolismo energtico. Entretanto, h grande variao na composio qumica da fibra de diferentes origens. Tal fato, associado s interaes que ocorrem entre alguns de seus constituintes, dificulta a avaliao do grau de utilizao de alimentos com elevados nveis de fibra para as espcies animais. Sendo assim, a preciso na anlise da composio qumica da frao fibrosa ao alimento ou da dieta deve ser a mais acurada possvel, para a correta estimativa das pores degradveis nos diferentes stios do trato gastrointestinal. A taxa de fermentao dos polissacardeos estruturais parece no estar associada a fatores isolados, no entanto, nos estudos em animais no-ruminantes tm-se relevado
50
que a natureza qumica da fibra, nvel diettico dos componentes fibrosos, forma de apresentao do alimento fibroso, grau de moagem da fonte de fibra e estado fisiolgico do animal, como as variveis mais importantes a serem consideradas. 4. ASPECTOS QUANTITATIVOS DA DIGESTO DOS CARBOIDRATOS. Quando o alimento chega ao segmento final do intestino delgado a digesto do amido e dos acares mais simples est quase completa. A literatura relata que os valores de digestibilidade do amido at este local do trato gastrointestinal variam entre 80 e 99%. Nos no ruminantes o grau de utilizao digestiva dos carboidratos pode variar dependendo de fatores tais como o prprio tipo de carboidrato, sua estrutura fsicoqumica, a interaes deste com outros componentes da dieta e a evoluo da atividade enzimtica com a idade. 4.1. Tipo de carboidrato. A utilizao digestiva dos carboidratos das plantas pelos no ruminantes melhor compreendida se levamos em considerao a sua acessibilidade ao enzimtica do animal ou dos microorganismos; para tanto, podemos distinguir trs tipos de grupos de carboidratos: 4.1.1. Grupo um: acares. De forma geral os mono e dissacardeos apresentam total disponibilidade no intestino delgado dos no ruminantes; contudo, caso que algum deles escape absoro at o leo terminal, sofrer a fermentao no intestino grosso e ceco. Os oligossacardeos rafinose e estaquiose so pouco degradados pelas enzimas prprias do trato digestivo dos no ruminantes mais so facilmente fermentveis no seu intestino grosso. 4.1.2. Grupo dois: polissacardeos de reserva. Em no ruminantes, a maioria dos amidos crus so hidrolisados no intestino delgado e absorvidos como glicose e a pequena parcela destes que resistente ao digestiva das enzimas prprias do trato fermentada no intestino grosso. A extenso com que o amido digerido, depende do mtodo de preparao do alimento. Assim, por exemplo, o amido dos cereais no estar plenamente disponvel para o animal ou para os microorganismos, se os gros no forem adequadamente quebrados, seja no processamento ou seja durante a mastigao. Contudo, importante levar em considerao que o grau de moagem dos gros depende tambm da espcie a que se destinam; no caso dos herbvoros no recomendvel a moagem fina destes. Uma vez alcanado o padro enzimtico de animal adulto, a digestibilidade dos hidratos de carbono, e fundamentalmente do amido, varia segundo a sua estrutura fsicoqumica. � Tamanho dos grnulos de amido. Quando comparados com os tubrculos, os cereais apresentam menor tamanho de grnulos o que parece explicar as diferenas na utilizao digestiva do amido entre estes dois alimentos.
51
As pesquisas realizadas mostraram que a digestibilidade ileal e fecal da matria orgnica caia quando em uma dieta controle era substituda 50 e 100% da cevada por batata Considerando que existe uma correlao alta entre o coeficiente de digestibilidade da matria orgnica e a digestibilidade da energia, a cevada oferece maior proporo de energia assimilvel na forma de glicose a nvel de leo do que a batata. � Diferena entre a proporo de amilose e amilopectina nos grnulos do amido. As variadas propores de amilose e amilopectina nos grnulo dos cereais geram diferenas quanto a digesto de amido no intestino delgado dos no ruminantes: os resultados das pesquisas mostram que maior a digestibilidade ileal do amido de trigo e milho que a do trigo. Entretanto, a digestibilidade da amilose e amilopectina so similares. � Processamento fsico mediante a utilizao de calor e umidade. O processamento fsico de distintas matrias primas ricas em amido outro fator que pode variar sua utilizao digestiva. A capacidade de perder o adquirir gua gera na estrutura da molcula de amido a presena de duas pores claramente diferenciadas na microscopia eletrnica. Estas so: � Gel: poro solvel em gua e facilmente atacada pelas enzimas -amilase; � Cristal: poro mais resistente ao ataque destas enzimas que, portanto, necessita ser liberada do grnulo. Entanto o aquecimento em meio mido aumenta o nmero de molculas hidratada, o aquecimento seco aumenta o de cristais. A capacidade de se hidratar ou no d as diferenas de digestibilidade entre os amidos de origens diversas. Os amidos da mesma origem vo diferir quanto digestibilidade, se forem cozidos ou no, porque a proporo das fases se altera. A hidratao dos gros ou a macerao um processo antigo e amplamente usado que apresenta os mesmos resultados de cozimento. Com a utilizao do calor e umidade, consegue-se captar gua no interior do grnulo que desestabiliza as pontes de hidrognio. Estas pontes unem distintas cadeias de polmeros e, quando apresentam-se instveis, permitem a liberao de amilose e amilopectina de estrutura granular. Este fenmeno, conhecido como gelatinizao, resulta particularmente adequado nos casos onde a digesto do grnulo de amido problemtica, como o caso dos tubrculos. Isto explica porque quando se submete a batata a um processo de calor e umidade, o coeficiente de digestibilidade melhora em relao s dietas que contm amido de batatas sem tratar. � Interao com outros componentes da dieta. Sabe-se que a adio de fibra dieta pode deprimir a digesto de amido. Uma possvel causa seria a menor disponibilidade do tempo para a atuao das enzimas digestivas sobre os substratos especficos, j que a adio de fibra dieta provoca um aumento da velocidade do trnsito. As pesquisas realizadas mostra que os efeitos da adio de fibra sobre a digestibilidade do amido dependem do nvel de fibra adicionado dieta e da fonte
52
de fibra utilizada: para um mesmo nvel de fibra bruta na dieta, o coeficiente de digestibilidade em porcas adultas, variou de 21% (palha de cevada) a 85% (farinha de soja). 4.1.3. Grupo trs: polissacardeos estruturais. A celulose e as hemiceluloses, constituintes da parede celular, esto disponveis para o animal somente mediante a ao de microorganismos. A lignina, embora no seja propriamente um carboidrato, visto que encontrada em estreita associao com os carboidratos da parede celular, geralmente tratada simultaneamente. Sob condies in vitro a celulose no lignificada facilmente atacvel pela microflora celuloltica do trato digestivo dos herbvoros, mas quando a proporo de lignina aumenta em conseqncia do envelhecimento da planta, a digesto da celulose se v seriamente interferida. Duas hipteses tm sido utilizadas para explicar esta interferncia: � Visto que a lignina indigervel, gera um bloqueio que no permite o ataque enzimtico das clulas vegetais; � A lignina ao se ligar quimicamente com os outros nutrientes, torna-os indigestveis. As interaes atravs de pontes de hidrognio que so encontradas freqentemente entre as hemiceluloses e a celulose comprometem o processo digestivo das hemiceluloses. As ligaes de tipo (14) da celulose no so hidrolisadas pelas amilases. Como o sistema digestivo dos vertebrados no secreta nenhuma enzima capaz de hidrolisar a celulose, esta no pode ser digerida e suas unidades de D-glicose no so totalmente aproveitveis como alimento para a maioria dos organismos superiores. Somente os microorganismos presentes no rmen dos ruminantes ou aqueles presentes no ceco e clon dos monogstricos, produzem celulases que digerem as ligaes beta da celulose e de hemiceluloses. As interaes atravs de pontes de hidrognio que so encontradas freqentemente entre as hemiceluloses e a celulose comprometem o processo digestivo das hemiceluloses. Os no-ruminantes herbvoros aproveitam relativamente melhor as hemiceluloses do que a celulose. Uma possvel explicao para maior eficincia de utilizao das hemiceluloses em relao celulose se baseia na hiptese de que as ligaes arabinofuranosdicas se mostram sensveis acidez gstrica o que possivelmente expe os resduos de xilose dos arabinoxilanos digesto. Parece que o principal problema na utilizao das hemiceluloses seria ento a dificuldade da remoo das cadeias laterais da arabinose pelos microorganismos no processo de fermentao. Em ruminantes a eficincia de desdobramento das hemiceluloses se torna semelhante da celulose. A digesto tambm parece depender da remoo da celulose incrustada o que promoveria uma barreira ao enzimtica. As ligaes complexas das hemiceluloses com a lignina tambm diminuem a sua degradabilidade.
53
As pectinas diferem do amido pela posio axial da ligao no carbono 4, no sendo assim atacadas pelas -amilases, porm so suscetveis ao microbiana. A degradabilidade das pectinas pelas bactrias intestinais de no-ruminantes quase que completa, ainda que produzam uma marcante alterao no volume da excreta fecal. A lignina o fator primrio que pode limitar o potencial de digesto da parede celular onde est quimicamente ligada. A limitao da digesto da lignina pode dever-se funo fsica desta como substncia que favorece a rigidez da parede celular, as caractersticas de suas unies qumicas com os polissacardeos estruturais, tambm conhecida como complexo lignocelulsica (neste caso a digesto mais afetada pelo grau e extenso das unies lignina-polissacardeos do que pela quantidade de lignina presente na dieta), a inibio da atividade enzimtica, ou as interaes de todos estes fatores. A presena de compostos fenlicos ou polifenlicos nos alimentos ou formados na degradao parcial da lignina no trato digestivo parece ser uma possvel causa de inibio da atividade enzimtica. Apesar de que parece existir uma forte relao negativa entre a quantidade de lignina e a digestibilidade das forragens, pode-se considerar que na determinao da digestibilidade da parede celular a composio qumica da lignina e as diferenas entre os tipos de ligaes entre esta e os demais componentes da parede talvez sejam mais importante do que a prpria quantidade de lignina. Nas monocotilednias, mormente em gramneas, por exemplo, existem ligaes steres entre os grupos cidos da lignina e as cadeias de xilanos de elevado peso molecular, enquanto que nas dicotilednias, em especial as leguminosas, so mais freqentes as ligaes glicosdicas com os grupos lcoois da lignina; esta hiptese explicaria porque a lignina das gramneas parece ter um efeito negativo maior sobre a digesto da parede celular do que a lignina das leguminosas. Ainda que sejam aceitos os efeitos, diretos ou indiretos, da indigestibilidade da lignina sobre a digestibilidade dos demais nutrientes da dieta, alguns pesquisas realizadas, sobretudo em ruminantes, do valores razoavelmente altos para o coeficiente de digestibilidade da lignina. Inmeros pesquisadores sugeriram que alguns dos resultados de digestibilidade relatados na literatura devem ser analisados luz das diferenas na prpria composio e dos erros na estimao qumica da frao lignificada. 4.2. Evoluo das enzimas digestivas com a idade. Um dos fatores mas marcante sobre o grau de digesto do amido e dos trs principais dissacardeos (maltose, sacarose e lactose) no intestino delgado a evoluo da secreo enzimtica dos no ruminantes com a idade. Nos sunos pode-se observar que a lactase apresenta atividade enzimtica alta mesmo desde o nascimento, descendo a nveis seis vezes menores durante os sete dias seguintes, mantendo-se mais ou menos constante ou ligeiramente em descenso em perodos posteriores at chegar ao nvel do animal adulto. Isto pode explicar porque sunos adultos alimentados com lactose podem desenvolver diarria e mal estar gasoso devido a uma deficincia de lactase. Em certos grupos populacionais muitos mamferos adultos apresentam intolerncia ao leite por terem deficincia de lactase. Inmeras causas podem explicar esta deficincia: parece ser que em algumas circunstncias trata-se de um carter autossmico recessivo herdado, geralmente expresso na adolescncia ou no incio da fase adulta.
54
Em outras, esta deficincia pode ser produzida pelo baixo consumo de leite na idade adulta. Seja qual for a causa desta deficincia enzimtica, no adulto aps a ingesto do leite a lactose acumula-se na luz do intestino delgado levando ao aumento de lquidos neste local o que causaria distenso de abdominal, nuseas, clicas e diarria lquida. A -amilase pancretica apresenta uma evoluo inversa registrada para a lactase: em geral ao nascimento a -amilase pancretica tem baixa atividade enzimtica, mas se incrementa rapidamente depois da 4 semana de vida. De maneira semelhante o comportamento da atividade enzimtica no caso da maltase. Ao nascimento a situao da atividade das sacarases um tanto semelhante da amilase pancretica e a maltase, porm ainda mais lento o ritmo de aumento da sua atividade com a idade. A deficincia de algumas dissacaridases especficas no aparelho digestivo traz como resultados transtornos gastrointestinais muito srios. Se durante as primeiras semanas de vida os mamferos recebem grandes quantidades de sacarose, desenvolvem diarrias severas e podem morrer por insuficincia de sacarase. Apesar de aumentar com a idade a capacidade dos animais para digerirem hidratos de carbono, parece necessrio, entretanto, manter parte dos carboidratos da dieta como sendo de origem lctea nos animais desmamados de maneira precoce a fim de no afetar negativamente os rendimentos produtivos. Tem sido observado que, em leites desmamados aos 21 dias, a velocidade de crescimento era maior quando a dieta com milho e soja continha tambm soro de leite. 5. Propriedades fsico-qumicas dos componentes da parede celular no processo digestivo. As propriedades fsico-qumicas da parede celular dos alimentos so importantes para entender seus efeitos fisiolgicos globais nos animais e no homem. Entretanto, o desenvolvimento destes conceitos tm sido mais evidentes nos trabalhos com aplicao em nutrio humana. As propriedades fsico-qumicas dos componentes parietais dos vegetais se caracterizam por influir sobre o trnsito digestivo dos alimentos, a absoro dos nutrientes e a adsoro dos sais biliares e o metabolismo dos lipdios. Por sua vez, a estrutura fsica, sua grande escala de polimerizao e a associao entre as molculas so os fatores de maior importncia que determinam algumas das propriedades da parede celular. A variao na composio qumica da parede influenciam a capacidade de absorver gua, de intercmbio catinico e de adsoro de substncias da matriz da digesta, que so algumas das propriedades fsico-qumicas da parede celular. O teor de lignina, por exemplo, influencia estas propriedades, determinando o grau de fermentao da digesta. O contedo de hemiceluloses e, particularmente o de pectinas, est relacionado com a capacidade de reteno de gua, ou capacidade higroscpica da parede, a qual, por sua vez, pode influir sobre a digesto e absoro de outros nutrientes, explicar o trnsito da digesta, o volume e peso das fezes assim como seu grau de viscosidade. O efeito negativo da capacidade higroscpica da parede celular sobre a digesto e
55
absoro de outros nutrientes no intestino delgado pode ser causado pelo aumento da massa que se produz ao se absorver quantidades significativas de gua posto que a matriz de digesta formada pode proteger alguns nutrientes da ao enzimtica. Embora exeram estes efeitos negativos, as substncias pcticas apresentam degradao completa no intestino grosso fornecendo fraes disponveis para a flora microbiana. Por outro lado, no ruminantes a viscosidade da digesta contribui a que seja mais lento o trnsito do alimento, em especial na primeira parte do trato digestivo, devido possivelmente resistncia que possa ter a digesta em relao s contraes do intestino. A presena de parede celular nas dietas dos no ruminantes favorecem a manuteno da flora do trato gastrointestinal e exercem um importante efeito tampo determinado pelas trocas catinicas. O complexo ecossistema intestinal, onde se incluem as colnias anaerbicas, est formado por mais de 400 espcies de bactrias, com uma contagem mdia fecal, para o caso do homem, de 1010 1012 por mililitro. Em herbvoros no ruminantes adultos a populao microbiana ceco-clica predominante formada por bacterides gram negativos, no esporulados do gnero Bacillus sp, com uma concentrao que pode variar entre 109 e 3,9x1011 por grama de contedo cecal em coelhos at 5x109 nos eqinos. A atividade bacteriana ceco-clica se produz sobre o substrato que escapou da hidrlise cida e enzimtica no estmago e intestino delgado e sobre os produtos gerados pela secreo endgena (enzimas, mucopolissacardeos e clulas de descamaes da mucosa). Os componentes da parede celular vegetal constituem-se no principal substrato que chega ao intestino grosso, exercendo uma forte influncia sobre o aumento da massa bacteriana e sua atividade enzimtica, sem afetar os tipos de bactrias. A capacidade de troca catinica da parede celular, medida pela sua capacidade para ligar-se aos ons metlicos em sua superfcie, pode conduzir alteraes na absoro de elementos minerais, afetar a ligao dos microorganismos aos polissacardeos estruturais e, portanto, a sua taxa de digesto. Substncias complexas, particularmente as que possuem grupamentos cidos como o urnico (pectinas) e fenlico (lignina), ou resduos sulfatados, e outros constituintes da clula vegetal (fitatos, silicatos e oxalatos) tendo sido apontadas freqentemente como responsveis pela interferncia negativa com a absoro mineral porquanto podem ligar-se ao Mg, Ca, Zn e Fe. As pectinas e, possivelmente a lignina, apresentam ainda especial interesse por sua capacidade de adsoro de cidos biliares com uma possvel repercusso sobre o metabolismo dos lipdios, constituindo uma propriedade de interesse para os estudos em nutrio humana devido aos efeitos sobre a hipocolesterolemia. Entre as teorias mais acatadas est a que prope que a ligao dos cidos biliares s fraes da parede celular reduz a sua reabsoro ileal afetando sua circulao enteroheptica. O colesterol circulante passa a ser imediatamente mobilizado para atender a sntese de cidos biliares, diminuindo assim a concentrao srica do colesterol. Um outro mecanismo que pode afetar o teor de colesterol srico deve-se ao do cido propinico, derivado da fermentao da parede celular. Os estudos in vitro realizados com hepatcitos isolados tm demonstrado que a sntese de colesterol inibida pela
56
presena do cido propinico; no entanto, sob condies experimentais in vivo, esta experincia ainda no ficou clara. 6. Algumas observaes sobre o uso de alimentos fibrosos nos no-ruminantes. A estratgia de utilizao digestiva dos componentes da parede celular nos noruminantes (onvoros, carnvoros e herbvoros) varia consideravelmente de acordo com a peculiaridade morfo fisiolgica do trato digestivo de cada espcie o que revela a adaptao evolutiva desses animais para conseguir a utilizao eficaz dos alimentos vegetais. Em coelhos, a estratgia utilizada para compensar suas elevadas necessidades metablicas e o consumo de alimentos de baixo valor nutritivo e ricos em parede celular, reside em uma alta capacidade de ingesto de alimento acompanhada de um rpido trnsito da digesta, sobretudo ao que se refere frao mais fibrosa, de maneira que o processo digestivo se manifesta principalmente sobre os demais componentes menos fibrosos da dieta. A fisiologia digestiva dos coelhos est igualmente relacionada com um importante processo fermentativo de digesto microbiana sobre os resduos da digesta, principalmente no ceco. Os fenmenos de adaptao relacionados com este processo entre os quais se incluem a dualidade da excreo fecal, denominada cecotrofia, e a ingesto (coprofagia) do material diferenciado excretado (cecotrofos) formam parte de um complexo sistema digestivo onde os componentes da parede celular ingeridos jogam papel de grande significado nutricional. Em coelhos, a exceo das fontes de fibra pouco lignificadas (polpas de beterraba e de ctricos), que apresentam coeficientes de digestibilidade da fibra de 70%, os da fibra bruta e fibra em detergente cido dos demais alimentos so normalmente menores que 20%. Apesar do escasso valor nutritivo da fibra no coelho, necessrio de se pensar em utilizar uma combinao entre diferentes fontes de fibra, e, sobretudo, em aportar um nvel mnimo de fibra indigestvel nas dietas visando evitar transtornos digestivos que podem conduzir a diarrias e, em um nmero elevado de casos, a morte do animal, principalmente durante a fase de crescimento do animal. A literatura especializada recomenda aportar um nvel de 12% de fibra bruta indigestvel ou entre 19 e 22% de FDA (ajustado a um mximo de 15 a 16% de amido) na dieta para evitar transtornos digestivos em coelhos em crescimento. Os pesquisadores espanhis defendem a idia de que em coelhos a melhor maneira para se estimar a fibra indigestvel e predizer a concentrao de energia da dieta utilizar a FDA devido a sua forte correlao negativa com como parmetro. A estratgia utilizada nos eqinos tambm est baseada no aumento de consumo de forma a compensar sua menor eficincia em utilizar polissacardeos estruturais da parede celular. Contudo, entre os herbvoros no-ruminantes, os eqinos apresentam maior capacidade de digesto frao fibrosa; eles digerem a fibra com uma eficcia equivalente 70% com respeito aos ruminantes. A frao fibrosa no entanto, deve apresentar boa palatabilidade afim de no influenciar o consumo. Ao igual que nos coelhos, a frao FDA dos alimentos tem se mostrado um potente estimador da concentrao de energia nos alimentos fornecidos aos eqinos. A utilizao de alimentos com alto teor fibroso tambm tem sido estudada na produo de sunos, com especial nfase em fmeas gestantes, onde os melhores resultados se mostram mais evidentes e cujas necessidades energticas so relativamente baixas.
57
Diversos autores assinalam que a incluso de certos alimentos fibrosos, em geral os menos lignificados, melhoram a reduo de custos do plantel de reproduo e diminuem a mortalidade ao nascimento, alm de que, quando so oferecidos prximos ao parto, previnem a constipao e a sndrome-mastite-agalaxia (MMA). Quando comparadas com animais em crescimento, as fmeas sunas gestantes possuem maior habilidade em obter a energia a partir dos alimentos fibrosos; contudo, em se tratando de cereais e outros alimentos com teores baixos de fibra, nenhuma diferena observada entre estas duas categorias. O maior desenvolvimento do trato gastrointestinal e o estabelecimento de uma flora especializada favoreceriam a maior degradabilidade da fibra nos animais adultos. Desde as pesquisas realizadas na dcada dos oitenta, ficou estabelecido que os sunos tm grande capacidade de utilizao digestiva dos nutrientes e a energia da alfafa (Medicago sativa), que em terminao podem obter at 30% de seus requerimentos de energia lquida para mantena a partir dos cidos graxos volteis produzidos no intestino grosso e que possvel manter o comportamento reprodutivo normal das fmeas at mesmo quando so alimentadas com dietas contendo acima de 90% de farinha de alfafa. Pesquisas realizadas em fmeas adultas alimentadas durante trs gestaes sucessivas com dietas que continham de 50 a 96% de feno de alfafa mostraram que era possvel manter rendimentos satisfatrios. Sobre esta mesma linha de pesquisa j foram relatados resultados que mostram que com a adio de at 50% de feno de alfafa se podia manter rendimentos aceitveis tanto em gestao quanto lactao; porm, estes rendimentos decresciam quando era atingido 95% de incluso de feno de alfafa na dieta. A influncia do nvel de fibra e da sua origem se d marcadamente no aproveitamento energtico dessas fontes a partir da fermentao microbiana no intestino grosso. Pesquisas realizadas na dcada dos anos noventa mostraram que o suprimento de energia pela fermentao pode representar 28% da energia lquida necessria para a mantena desde que sejam includos na dieta nveis de 20% de alimentos fibrosos de alta degradabilidade como a polpa de beterraba. Pesquisadores espanhis defendem a incluso de fontes fibrosas fermentecveis como as polpas de beterraba e de polpa de ctricos em sunos adultos como uma alternativa para diminuir custos de produo sem afetar o rendimento produtivo dos animais. As pesquisas realizadas at o momento sobre o uso de recursos alimentcios fibrosos em sunos permitiram assinalar que existem efeitos associativos do nvel de incorporao da fibra nas dietas sobre o animal, desde a mesma ingesto de alimentos at os vrios processos que acontecem no trato gastrointestinal. Contudo, os estudos mais recentes tm sugerido que os efeitos fisiolgicos e nutricionais da fibra dependem no s do nvel da parede celular incorporada dieta, mas tambm de sua composio qumicas e estrutural, a forma como est fisicamente associada com outros nutrientes e o animal no que tem a ver com seu estado fisiolgico mas, principalmente, do local do trato gastrointestinal onde esto acontecendo os processos digestivos. Embora a incorporao de nveis crescentes de alimentos fibrosos s dietas de sunos fornecem uma fonte adicional de energia aos gros de cereais vista a contribuio da fermentao microbiana dos constituintes da parede celular no ceco e colo, os relatos das pesquisas salientam que existem efeitos associativos entre o nvel de utilizao e a
58
composio da fibra sobre a prpria ingesto de alimento, os processos digestivos, as caractersticas e a composio das fezes e, portanto, a digestibilidade dos nutrientes. Duas estratgias tm sido utilizadas para estudar os efeitos da fibra na alimentao de sunos. A primeira tem-se baseado na substituio parcial ou a suplementao de dietas semi-sintticas por alguns isolados de fibra (celulose, gomas e pectina). A outra estratgia apoia-se no estudo de alguns alimentos fibrosos propriamente ditos; nesta j foram pesquisadas palhas, cascas e resduos fibrosos da safra, subprodutos da transformao dos cereais e oleaginosas e derivados da indstria de extrao de sucos (polpa de ctricos) e da obteno de acar (polpa de beterraba). A escolha da primeira estratgia e no a dos alimentos fibrosos tem algumas vantagens porm est aberta crtica. Os efeitos dos dois tipos de fraes da parede celular (fibra solvel e insolvel) sobre a funo gastrointestinal esto altamente relacionados com as suas propriedades fsicas e qumicas (capacidade de absoro de gua, adsoro de minerais, tamponante, de troca catinica e de formao de gis) e com o local onde esto acontecendo os processos digestivos. No intestino delgado a fibra solvel tem seu maior impacto atravs da reduo da taxa de absoro de vrios nutrientes, sendo a mais marcante para a glicose, o colesterol plasmtico e os aminocidos, diminuindo, consequentemente, a digestibilidade ileal dos aminocidos, lpides e minerais. Este efeito pode ser explicado por uma reduo no vazamento gstrico, um efeito sobre a difuso e absoro dos nutrientes ou um aumento da viscosidade do contedo digestivo. Neste mesmo local do trato digestivo, a fibra solvel, entretanto, afeita a diluio do contedo ileal, diminui o tempo de passagem da digesta e aumenta o volume fecal. No intestino grosso pode ser dito que, de maneira geral, a fibra da dieta muda a atividade das bactrias e, em conseqncia, altera o metabolismo do nitrognio, seus padres de excreo e, possivelmente, afete o balano geral de nitrognio do animal; porm, as causas que explicam a origem do aumento do volume fecal e das perdas do nitrognio nas fezes tambm dependem do tipo de fibra que atingi este local do trato digestivo: no caso da fibra solvel, por exemplo, explicado pelo aumento na excreo do nitrognio microbial; no entanto, a frao insolvel, visto que apresenta baixa degradabilidade aumentam a excreo da parede celular ligada protena explicando, assim, o maior volume fecal e o aumento na excreo do nitrognio nas fezes. O efeito global de ambos os dois mecanismos traduz-se em uma diminuio da digestibilidade aparente do nitrognio. Em aves, a contribuio positiva da fibra considerada incerta devido a alta indigestibilidade dos componentes da parede celular. As pesquisas realizadas demonstraram que os complexos celulsicos, as hemiceluloses e as substncias pcticas so pouco digeridos mesmo pelas aves adultas. As mesmas pesquisas tambm tm mostrado que dos polmeros estruturais observa-se alguma degradao das pentosanas solveis em gua presentes em algumas hemiceluloses, particularmente as encontradas em gros de cereais como o trigo. Os efeitos dos NPS ou polissacardeos no-amilceos das plantas (celulose, arabinoxilana, arabinogalactana, galactomanana, xiloglucana e substncias pcticas)
59
sobre a digesto tm sido estudados recentemente em frangos de corte. A figura 2 apresenta um resumo dos principais achados nesta rea. De uma forma geral, o aumento nos contedos de fibra na dieta para aves diminui os valores de EM da mesma: em poedeiras pode-se considerar que por cada aumento de um ponto na fibra bruta h uma queda de 90 kcal de EM/kg; caso que o estimador utilizado seja a FDA a queda de 80 kcal de EM/kg e de 40 kcal/kg quando o estimador usado a FDN. Nas aves, a frao fibrosa, no en tanto, revela-se como um bom preditor do valor nutricional dos alimentos, devido alta correlao negativa que existe entre o seu contedo e a digestibilidade da protena, gorduras, matria orgnica e EM. As informaes disponveis para ces indicam que 5% o nvel timo de fibra para ser includo nas dietas, visto que este promove a normalizao das funes gastrointestinais, j nveis dietticos entre 10 e 15% de fibra seriam para atender o manejo alimentar de animais obesos o diabticos; nveis acima de 15% de fibra bruta na matria seca ingerida podem deprimir o trnsito digestivo aumentando, porm, de maneira significativa o volume fecal e o nmero de defecaes dirias. Efeitos positivos da fibra sobre a saciedade em ces que necessitam estar sob restrio calrica tambm tm sido relatados na literatura. V. METABOLISMO DOS CARBOIDRATOS. No aparelho digestivo, somente os monossacardeos se absorvem, com exceo nos animais recm-nascidos que podem absorver molculas mais volumosas. Por conseguinte, para que a absoro se realize, as enzimas digestivas ou a flora microscpica presentes no intestino grosso devero hidrolisar os poli, tri e dissacardeos. As carboidrases so efetivas na hidrlise da maioria dos carboidratos complexos a monossacardeos, com exceo daqueles que possuem o enlace Glicose -Glicose, tal como a celulose. 1. ABSORO. Encontram-se no duodeno e no jejuno os principais stios de absoro de monossacardeos. Na poro do leo inferior, no estmago e no intestino grosso se absorvem poucos aucares. A velocidade de absoro varia segundo o tipo de monossacardeo. A glicose e galactose so absorvidas mais rapidamente do que a frutose; os demais monossacardeos so absorvidos em uma velocidade todavia menor. Estas diferenas se devem existncia, para os primeiros, de mecanismos especficos de transporte ativo, enquanto que para a frutose necessrio a converso prvia da glicose. O resto de monossacardeos so absorvidos por difuso simples. A converso de alguns monossacardeos em glicose se produz dentro da clula da mucosa intestinal. Os monossacardeos que no se convertem em glicose, neste local durante a absoro, podem se converter em glicose atravs de reaes no fgado, onde
60
sero utilizados nas distintas rotas metablicas para a obteno de energia, formao de gordura, etc. Os cidos graxos volteis produzidos durante a fermentao dos materiais fibrosos so cidos de cadeia curta e, por isso mesmo, solveis em gua. Na sua maior parte, so absorvidos no prprio local da digesto microbiana e utilizados metabolicamente para finalidade semelhante glicose, porm com uma eficcia menor, em torno de 75%. A importncia quantitativa destes processos varia amplamente. Assim segundo vrios autores, os cidos graxos volteis produzidos no intestino grosso de no-ruminantes podem cobrir entre 5 e 30% das necessidades da energia lquida de mantena. Por outro lado, a eficcia de utilizao da energia que se absorve no ceco a metade da que se absorve ao nvel do intestino delgado. 2. METABOLISMO DA GLICOSE. A glicose pode ser utilizada pelo mecanismo animal de duas formas: � Mediante via anablica: sntese de glicognio, lipdeos e lactose. � Mediante via catablica: gliclise e Ciclo de Krebs. 2.1. Gliconeognese e Gliclise. O organismo animal armazena pouqussima energia sob a forma de carboidrato, mas parte da glicose se converte em glicognio, o qual armazenado no fgado e msculo esqueltico. O glicognio um composto similar ao amido que pode converter-se novamente, de forma rpida, em glicose. O nvel do acar sangneo se mantm dentro de uma margem muito estreita em animais normais. Ao converter a glicose sangnea circulante em glicognio (gliconeognese), o organismo volta a convert-lo em glicose por meio da gliclise; quando descendem os nveis sangneos. A concentrao de glicose sangnea se eleva depois da ingesto de alimentos, mas regressa aos nveis de jejum em poucas horas. Esta homeostase se controla de forma endcrina; os hormnios so importantes na manuteno da concentrao de glicose no sangue dentro dos limites normais das espcies. O armazenamento de glicognio depois da ingesto de alimentos evita a elevao do acar sangneo (hiperglicemia). A formao do glicognio limitada. Quando a ingesto de carboidratos excede as necessidades de armazenamento de glicognio, a glicose se converte em gordura. 2.2. Gliconeognese. A principal rota da glicose na maioria dos tecidos comea com a fosforilao da glicose glicose-6 fosfato; esta se transforma em glicose-1-fosfato e polimeriza-se em glicognio. Uma pequena quantidade de glicose-6-fosfato entra continuamente em outra rota, chamada de Via das Pentoses. Esta via primariamente importante como fonte de ribose para a sntese de cidos nuclecos (RNA e DNA) e nucleotdeos enzimticos. A maior rota da glicose-6-fosfato a via glicoltica que, aps a transformao da glicose-6-fosfato e, vrias reaes, produz piruvato. O piruvato marca o segundo ponto importante no metabolismo da glicose. Ele permanece no citoplasma e se transforma, anaerobicamente, em lactato ou entra na mitocndria e se transforma em Acetil- CoA. Este composto qumico representa um ponto bsico dentro de grande nmero de vias
61
metablicas existente no organismo animal, pois ele pode resultar da degradao de cidos graxos volteis, glicose, aminocidos e lipdeos. Na mitocndria, o piruvato passa para outra via metablica, chamada de Ciclo de Krebs, Ciclo do cido tricarboxlico ou, Ciclo do cido cctrico. O Ciclo de Krebs o final das vias metablicas energticas comum aos esqueletos carbonados que deram origem Acetil-CoA. H pequenos rumos metablicos ao longo do Ciclo de Krebs que so importantes nas interconverses, como a transformao de glicose e sntese de cidos graxos e lipdeos. O resultado final do Ciclo de Krebs a oxidao do carbono e hidrognio para a formao de duas molculas de adenosina trifosfato (ATP). O ATP um componente rico em energia qumica. utilizado nos processos bioqumicos que requerem energia para trabalho qumico (biossntese), para trabalho mecnico (contrao muscular) e trabalho osmtico (transporte ativo). 3. Metabolismo dos cidos graxos volteis. Os cidos graxos volteis podem ser utilizados em processos catablicos ou anablicos. No catabolismo, seu destino a sntese de ATP. No anabolismo, so empregados na sntese de lipdeos, e em particular no caso do cido propinico, na sntese de glicose. O cido actico metabolizado no tecido perifrico (tecido muscular e adiposo). Entra no ciclo de krebs como Acetil-CoA. utilizado na sntese de cidos graxos de cadeia longa e para cada molcula de cido actico so produzidos 10 moles de ATP. O cido propinico metabolizado no fgado onde transforma-se em glicose ou metabolizado via Ciclo de Krebs para a produo de 18 moles de ATP por molcula de cido oxidado. O cido butrico no fgado convertido Acetil-CoA que entra no Ciclo de Krebs e produz 27 moles de ATP por molcula de cido. VI. BIBLIOGRAFIA DE APOIO. Andrigueto, J.M. et al. Nutrio Animal. So Paulo, Nobel. Ed. da Universidade Federal do Paran, 1982. Bergman, E.N. Contributions of VFA from the gastrointestinal tract in various species. Physiological Reviews. 70 (2): 567-590. 1990. Champ, M. Digestion des glucidez chez le monogastrique. reproduction, Nutrition and Dveloppment. 25 (4B): 819-842. 1985. Corring, T.; Juste, C.; Lhoste, E.F. Nutritional regulation of pancreatic and biliary secretions. Nutrition Research Review. 2: 161-180. 1980.
62
Cranwell, P.D. Development of the neonatal gut and enzyme systems. p 99-154. In: Varley, M.A. The neonatal pig. Development and survival. CAB International. 1995. Drochner, W. Digestion of carbohydrates in pig. Archives of Animal Nutrition. 43 (2): 95-116. 1992. Ferreira, M.W. Os componentes da parede celular vegetal na nutrio de noruminantes. P. 85-113 In: Simpsio Internacional de Produo de NoRuminantes. Anais da XXXI Reunio Anual da Sociedade Brasileira de Zootecnia. 17-21 julho de 1994. Maring.Paran. Hintz, H.F.; Cymbaluk, N.F. Nutrition of horse. Annual Review of Nutrition. 14:243268. 1994. Johnson, L.R. Regulation of gastrointestinal mucosal growth. Physiological Reviews. 68 (2): 456-502. 1988. Lassiter, J.W. & Edwards JR.,H.M. Animal Nutrition. Reston, Virgnia. Reston Publishing Company, Inc. 1982. Low, A.G. Nutritional regulation of gastric secretion, digestion and emptying. Nutrition Research Review. 3: 229-252. 1990. Marounek, M. et al. Distribution of activity of hidrolytic enzymes in the digestive tract of rabbits. Britishy Journal of Nutrition. 73 (3): 463-469. 1995. Marty, J.; Vernay, M. Absorption and metabolism of the VFA in the hind-gut of the rabbit. British Journal of Nutrition. 51: 265-278. 1984. Maxwell, F.J.; Stewart, C.S. Microbiology of the gut and the role of probiotics. p. 155-186. In: Varley, M.A. The neonatal pig. Development and survival. CAB International. 1995. Maynard, L.A. & Loosli, J.K. Nutrio Animal. Rio de Janeiro. Freitas Bastos. 1974. McDonald, P. et al. Animal Nutrition. London, Longman London and New York, 1973. Mei, N. Intestinal chemosensitivity. Physiological Reviews. 65 (2): 211-237. 1985. Nabuurs, M.J.A. Microbiological, structural and functional changes of the small intestine of pigs at weaning. Pigs News nad Information. 16 (3): 93N-97N. 1995. Nitsan, Z. et al. Growth and development of the digestive organs and some enzymes in broiler chicks after hatching. British Poultry Science. 32: 515-523. 1991. Perry, T.W. Animal Life-Cycle Feeding and Nutrition. Orlando, Flrida. Academic Press, Inc. 1984.
63
Pinchasov, Y. Early transition of the digestive system to exogenous nutrition in domestic post-hatch birds. British Journal of Nutrition. 73 (3): 471-478. 1995. Pluske, J.R. et al. Nutrition of the neonatal pig. p. 187-238. In: Varley, M.A. The neonatal pig. Development and survival. CAB International. 1995. Potty, V.H. Physio-chemical aspects, physiological functions, nutritional importance and technological significance of dietary fibres- A critical appraisal. Journal Food Science Technology. 33 (1): 1-18. 1996. Rrat, A. et al. Absorption kinetics of some carbohydrates in conscious pigs. Qualitative aspects. British Journal of Nutrition. 51: 505-516. 1984. Quantitative aspects. British Journal of Nutrition.. 51: 517-529. 1984. Smits, C.H.M.; Annison, G. Non-starch plant polysaccharides in brolier nutritionTowards a physiologically vallid approach to their determination. Worlds Poultry Science Journal. 52: 203-221. 1996. Yamauchi, K.E.; Isshiki, Y. Scanning electron microscopic observations on the intestinal villi in growing white leghorn and broiler chickens from 1 to 30 days of the age. British Poultry Science. 32: 67-78. 1991. DIGESTO E METABOLISMO DOS LIPDEOS SUMRIO 1 Introduo 2 Funes principais dos lipdeos 3 Definio de lipdeos 4 cidos graxos 4.1.1 Estrutura 4.1.2 Propriedades fsicas e qumicas 4.1.3 Ocorrncia 5 Classificao dos lipdeos 6 Metabolismo de lipdeos 6.1.1 cidos graxos 6.1.1.1 Biossntese 6.1.1.2 Catabolismo 6.1.2 Triacilgliceris 6.1.2.1 Biossntese 6.1.2.2 Degradao 7 Digesto, absoro e utilizao 7.1 Digesto 7.1.1 Digesto pr-duodenal 7.1.2 Digesto duodenal 7.2 Absoro 7.3 Utilizao 7.3.1 Digestibilidade 7.3.2 Deposio de gordura 7.3.3 Mobilizao de reservas
64
8 Literatura consultada 1 INTRODUO Os lipdeos, ou gorduras, apresentam um problema digestivo especial para o animal, porque no se dissolvem em gua, o principal meio pelo qual ocorre a maioria dos processos corpreos, incluindo a digesto. A ao detergente necessria para emulsificar ou dissolver lipdeos, de forma que possam ser submetidos s aes das enzimas hidrolticas, hidrossolveis, no intestino. O problema da solubilidade torna a mecnica da digesto e da absoro dos lipdeos ligeiramente diferentes daquela das protenas e carboidratos. Os lipdeos compem uma grande poro das dietas dos carnvoros, ao passo que em geral formam uma poro menor das dietas naturais dos herbvoros adultos. Apesar disso, parece que as espcies herbvoras possuem a capacidade de digerir e absorver lipdeos em quantidades considervelmente mais elevadas do que as encontradas em suas dietas naturais, e freqentemente, lipdeos suplementares so acrescidos s dietas de cavalos de corrida e vacas leiteiras de alta produo. Os neonatos de todas as espcies mamferas possuem alta capacidade de digesto e absoro de lipdeos, porque o leite tem alto teor de gordura. O lipdeo diettico primrio o triacilglicerol (TG), que tanto pode originar-se de fontes vegetais como animais. Outros lipdeos dietticos importantes incluem o colesterol e o ster de colesterol de origem animal, ceras de fontes vegetais e fosfolipdeos, tanto de fontes vegetais como animais. A estrutura desses vrios elementos podem ser visualizadas nas figuras 1, 2 e 3. 2 FUNES PRINCIPAIS DOS LIPDEOS As funes gerais dos lipdeos so as seguintes: 1 Fornecimento de energia para manuteno normal do organismo e das funes produtivas. A hidrlise total dos TG proporciona glicerol e cidos graxos (AG) que servem como fontes concentradas de energia (2,25 vezes mais energia que carboidratos e protenas) 2 Fonte de AG essenciais : os animais aparentemente no sintetizam o cido linoilico e o cido linolnico, ou pelo menos no em quantidades suficientes para prevenir as alteraes patolgicas e, portanto, devem estar presentes nas dietas. Na prtica, cido linolico adicionado dieta destinada s aves, todavia na alimentao de sunos esse AG no precisa ser adicionado como suplemento, pois todos os alimentos so relativamente ricos em AG essenciais. No freqente o aparecimento de deficincia de AG essenciais ( diminuio da velocidade de crescimento, alopecia, dermatite e problemas reprodutivos). O araquidnico sintetizado a partir do linolico. Ento diz-se que os cidos linolico e linolnico so nutricionalmente essenciais, enquanto o cido araquidnico metabolicamente essencial.
65
A importncia metablica dos AG essenciais parece derivar de suas funes especficas: - Papel estrutural como constituintes das membranas celulares: dietas deficitrias em AG essenciais implicam menor permeabilidade e, portanto, reduo do intercmbio de nutrientes da clula. - Papel como precursores na biossntese de substncias tais como prostaglandinas ( sintetizadas a partir do araquidnico), prostaciclinas, troboxanos e leucotrienos que atuam como reguladores metablicos de importantes processos como funcionamento do rim, reproduo, regulao da presso sangunea, coagulao do sangue, resposta imunolgica etc. - Portador das vitaminas lipossolveis: a absoro das vitaminas lipossolveis funo da digesto e absoro das gorduras. As vitaminas lipossolveis encontram-se na luz do tubo intestinal, dispersas em micelas parecidas ou idnticas quelas formadas por ocasio da absoro de AG. As micelas mistas que contm monoacilgliceris (MG) e AG livres captam as vitaminas lipossolveis de maneira mais eficiente que as micelas que no contm esses compostos. 3 DEFINIO DE LIPDEOS Lipdeos so substncias orgnicas oleosas ou gordurosas, insolveis em gua, extradas das clulas e tecidos por solventes orgnicos no polares como o clorofrmio e o ter. Os lipdeos mais abundantes so as gorduras ou TG, que so os principais combustveis na maioria dos organismos, sendo a principal forma de armazenamento de energia qumica. Os lipdeos so estruturalmente bastante diversificados, no entanto apresentam um aspecto qumico comum: todos so derivados do acetato.
4 CIDOS GRAXOS 4.1 ESTRUTURA So os principais componentes dos lipdeos, aos quais conferem suas propriedades gerais. A estrutura geral R-COOH, onde R equivale a uma cadeia de carbonos, que contm desde 2 at 24 ou mais tomos. Sua caracterstica fundamental possuir uma funo cida de natureza carboxlica hidrfila e uma cadeia parafnica hidrfoba. R-COOH Cadeia parafnica Hidrfoba carboxila hidrfila
cido butrico CH3 (CH2)2COOH
66
H H H O H C C C C OH H H H H Cada tomo de carbono da cadeia, com exceo do grupo carboxila e do grupo metil terminais, tem dois tomos de hidrognio aderidos a ele. cido linolico (n-6) CH3(CH2)4-CH=CH-CH2-CH=CH- (CH2)7 COOH A cadeia parafnica pode ser saturada (somente ligao simples entre carbonos) ou insaturada ( uma ou mais ligaes duplas entre carbonos). Nas cadeias poliinsaturadas, um tomo de hidrognio eliminado nas ligaes duplas.. A desidrogenao torna o princpio nutritivo lipdeo mais digestvel. Ao contrrio, as gorduras mais hidrogenadas ou saturadas, apresentam menor digestibilidade. As cadeias parafnicas podem ser linear, ciclica ou ramificada. Distinguem-se, ainda, trs sries de AG insaturados: n-9, n-6 e n-3 conforme a ligao dupla esteja no nono, sexto e terceiro carbono desde o grupo metil do cido graxo, respectivamente (figura 6). 4.1.2 PROPRIEDADES FSICAS E QUMICAS O ponto de fuso mais elevado nos AG saturados (que so slidos a temperatura ambiente), aumentando com o tamanho da cadeia desde 44,2C no cido lurico (12:0) at 76,5C no cido araqudico (20:0). O ponto de fuso diminue pela introduo de duplas ligaes, assim passa de 69,6C no cido esterico (18:0); 13,4C no cido olico (18:1); -5C no linolico (18:2) at -11C no cido linolnico (18:3). Os AG de cadeia longa, da mesma forma que seus steres com glicerol, so inodoros a temperatura ambiente; os AG de cadeia curta tm, entretanto, um cheiro desagradvel, assim como alguns de seus principais derivados da rancificao dos AG. 4.1.3 OCORRNCIA O contedo em AG varia de um alimento a outro. A maior parte dos leos vegetais (milho, soja, girassol, algodo, amendoim, colza) se caracterizam por um alto contedo em cido olico e linolico (tabela 1); alguns destes leos apresentam particularidades como aprecivel nvel de linolnico no leo de soja; ercico (22:1) em algumas variedades de colza e malvlico e esterclico (AG ciclopropenides 18:1 e 19:1 respectivamente) no leo de algodo. Tabela 1. Composio em AG (% peso) de diferentes leos vegetais cidos graxos1 8:0 10:0 leo girassol leo de soja Gord. De coco 6,0 7,0 leo palma -
67
12:0 48,0 14:0 0,4 18,0 2,0 16:0 4,0 9,0 9,0 36,0 18:0 2,0 2,0 2,0 5,0 18:1 33,0 20,0 8,0 47,0 18:2 60,0 54,6 1,0 10,0 18:3 12,0 1 o primeiro nmero indica o comprimento da cadeia (nmero de tomos de carbono) e o segundo as duplas ligaes Outros leos vegetais como o de coco e palmiste caracterizam-se por seu alto contedo em AG de cadeia mdia (especialmente lurico), enquanto que no leo de palma destaca-se a concentrao em cido palmtico (16:0) e olico (tabela 1). As gorduras animais diferem marcadamente da composio mdia dos leos vegetais por seu maior grau de saturao, sendo os AG predominantes no tecido adiposo o palmtico, esterico e olico (tabela 2). Dentro deste grupo, a gordura do gado suno (manteiga) ou de aves so mais insaturadas que as de ruminantes (sebo).
Tabela 2. Principais AG dos triacilgliceris do tecido adiposo, em diferentes espcies animais cidos graxos 14:0 16:0 16:1 18:0 18:1 18:2 Bovino 3,7 21,8 4,7 17,1 42,3 2,3 Composio (% peso) Ovino 2,9 23,7 3,5 18,3 43,2 3,8 Suno 1,5 27,6 3,2 12,2 45,1 10,4
A composio em AG da gordura do leite varia entre espcies refletindo diferenas na sua fisiologia digestiva. Assim, a gordura do leite de vaca tem elevada percentagem de AG de cadeias curta e mdia, procedentes do metabolismo dos cidos graxos volteis do rmen, enquanto que a de porca similar a gordura do tecido adiposo (tabela 3). Os leos de pescado distinguem-se dos dois grupos anteriores por maior contedo de AG de cadeia longa (com mais de 18C) da famlia do cido linolnico (srie n-3), com alto grau de insaturao (tabela 4). Tabela 3. Composio em AG (%) do leite de vaca e de porca
68
AG 4:0 6:0 8:0 10:0 12:0 14:0 16:0 16:1 18:0 18:1 18:2
Leite de vaca 3,3 2,5 1,7 4,2 5,8 15,3 39,4 2,7 3,5 14,4 1,1
Leite de porca 4,5 35,4 11,1 3,9 34,7 10,5
Tabela 4. Composio em AG (% peso) de dois tipos de leo de pescado cidos graxos 14:0 16:0 18:0 16:1 18:1 20:1 22:1 Total n-6 Total n-3 A. LIPDEOS SIMPLES So steres de cidos graxos com certos lcoois, particularmente o glicerol, o colesterol e o cetlico. As gorduras e leos so steres de AG com glicerol e as ceras so steres de AG com lcoois diferentes do glicerol. A.1 TRIACILGLICERIS (TG) So os lipdeos mais abundantes (mas de 90% do total) nos alimentos utilizados habitualmente em alimentao animal e tambm o so na gordura do tecido adiposo e na das produes animais. So steres de glicerol com AG (figura 1). As principais variaes de seu valor nutritivo esto ligadas a diferenas no tipo de AG que formam parte de sua molcula. J foram isolados mais de 100 AG distintos dos TG de diversas clulas e tecidos. Os mais comuns tm um nmero par de tomos de carbonos e uma cadeia linear e diferem no tamanho da cadeia (entre 12 e 22 tomos de carbono) e em seu grau de insaturao, como mostrado na figura 4. So os componentes principais de armazenamento ou depsito de gorduras nas clulas vegetais e animais, mas no so normalmente encontrados nas membranas. Nutricionalmente este o grupo mais importante, dado o seu uso como ingredientes nas dietas. As gorduras e os leos presentes na maioria das substncias comestveis, Anchoveta 7,5 17,5 4,0 9,0 11,6 1,6 1,2 2,1 33,7 5 CLASSIFICAO DOS LIPDEOS Arenque 6,1 10,8 1,4 7,3 10,3 13,4 21,3 1,3 21,4
69
caracterizam-se pelo seu alto valor energtico. Um grama de gordura comum produz aproximadamente 9,45 cal, quando se submete a combusto total, comparado a 4,1 cal que produz um carboidrato comum ou 4,4 cal de uma protena. Assim a gordura produz aproximadamente 2,25 mais energia que carboidrato e protena. Anlises relacionando a proporo dos AG constituintes das gorduras indicam o mximo de energia que pode ser esperado. A energia bruta dos AG saturados aumenta com o comprimento da cadeia porque as unidades - CH2 no oxidadas, progressivamente ocupam uma proporo maior da molcula em relao ao grupo carboxila totalmente oxidado. Possuir uma ligao insaturada na cadeia um passo oxidativo parcial, e o contedo de energia bruta diminui proporcionalmente. A . 2 ESTERIDES So lipdeos no saponificveis, isto , no so hidrolizveis pelo aquecimento com lcalis, para produzir sabes com seu componente AG. Os esterides mais abundantes so os esteris, que so lcoois esterides (figura 3). O colesterol o principal esterol nos tecidos animais. O colesterol e seus steres com AG so componentes importantes das lipoprotenas plasmticas e da membrana celular externa. So, ainda, precursores das vitaminas D2 e D3 , de hormnios sexuais e de sais biliares. A molcula de colesterol apresenta um grupo cabea polar e uma estrutura no polar no hidrofbica (quatro anis rgidos). A . 3 CERAS E CERDEOS So steres de AG com lcoois de cadeia longa. Os AG possuem cadeias longas (25 a 35C) saturadas e insaturadas (figura 3). Atuam como revestimento protetor natural das folhas, frutos, caules, insetos, pele, penas, pelos e servem como material estrutural das colmeias. So, tambm, os principais lipdeos alimentares de reserva na cadeia ocenica. No so usados nutricionalmente, por serem altamente hidrfobos, no sendo atacados por enzimas de animais superiores. Podem ser usados como protetores de substncias que no devem ser degradas pelo estmago. No duodeno, emulsificam-se com os sais biliares. B. LIPDEOS COMPOSTOS So steres de AG , que contm outros grupos alm de um lcool e um AG, tais como fosfolipdeos, glicolipdeos e lipoprotenas. B.1 FOSFOLIPDEOS Tm uma estrutura qumica similar a dos TG, com a particularidade de que um dos grupos hidroxlicos do glicerol est esterificado com cido fosfrico e este, por sua vez, com um radical nitrogenado. Este tipo de estrutura confere molcula um alto grau de polaridade do que deriva o importante papel que tm essas substncias na digesto das gorduras.
70
Encontram-se principalmente na bile e nas membranas celulares (tanto animais como vegetais), estando sua estrutura relacionada com a regulao da permeabilidade da membrana, com o transporte, absoro e metabolismo de AG, do sdio e do potssio, com as oxidaes celulares, com a coagulao do sangue e com a reserva de AG e fosfatos. Por outro lado, como se observa na tabela 5, os cidos dos fosfolipdeos do tecido muscular tm um grau de insaturao notavelmente superior a dos TG do tecido adiposo. Tabela 5. Composio (% em peso) em AG dos fosfolipdeos do msculo longissimus dorsi em diferentes espcies animais AG 14:0 16:0 16:1 18:0 18:1 18:2 18:3 20:4 Bovino 0,4 22,6 2,5 7,8 24,3 23,0 2,0 12,5 Ovino 2,1 22,0 2,3 13,2 30,3 18,0 3,9 Suno 0,2 18,9 1,6 12,0 18,8 25,5 0,2 7,7
O fosfolipdeo mais comum a lecitina que contm colina como radical nitrogenado; as lecitinas obtm-se comercialmente como subprodutos da fabricao de leos vegetais e podem ser utilizadas como aditivos em alguns tipos de dieta. B.2 LIPOPROTEINAS As lipoprotenas contm tanto lipdeos polares e TG como o colesterol e seus steres. So formas de transporte de lipdeos na linfa e no plasma sangneo. Esto presentes em todos os glbulos de gordura da gema do ovo, leite etc. H trs classes principais de lipoprotenas, de acordo com seu contedo lipdico, isto , sua densidade. O plasma sanguneo tambm contm quilomcrons(figura 5), estruturas muito maiores que as lipoproteinas, mas com densidade muito baixa (transportam triacilgliceris do intestino delgado, onde so absorvidos durante a digesto, at os depsitos de gordura). B.3 GLICOLIPDEOS So glicerdeos que apresentam um carboidrato em substituio a um AG (nas posies 1 ou 3). Com frequncia os glicolipdeos tm em sua composio nitrognio. Juntamente com os fosfolipdeos e steres de colesterol, compem a membrana plasmtica. C. LIPDEOS DERIVADOS Incluem substncias que derivam dos grupos mencionados anteriormente atravs de uma hidrlise como AG, glicerol e outros lcoois.
71
D. TERPENIDES So caracterizados por uma unidade que se repete, o isopreno H2C = C C = CH3
CH2
Neste grupo incluem-se os carotenides, xantofilas, tocoferis e as vitaminas A e K. 6. 6 METABOLISMO DE CIDOS GRAXOS E TRIACILIGLICERIS Os AG e os TG so utilizados na deposio de tecido adiposo e da mobilizados como fonte de energia ou para sntese de lipdeos corporais (leite, ovos etc). A glndula mamria, o fgado e o tecido adiposo so os principais lugares onde se processa a biossntese de AG e TG. O fgado o rgo central para a interconverso e o metabolismo de lipdeos (figura 7) e sua funo pode resumir-se em processar e distribuir lipdeos (e tambm outros nutrientes). Assim, resumidamente, pode-se enumerar as seguintes atividades metablicas do tecido heptico: sntese de AG a partir de carboidratos, AGV e lipdeos degradados sntese de AG a partir de aminocios lipognicos sntese de colesterol a partir de acetil CoA sntese de fosfolipdeos e lipoproteinas sntese de corpos cetnicos degradao de AG e fosfolipdeos remoo de fosfolipdeos e colesterol da corrente sangunea alongamento e encurtamento de cidos graxos desidrogenao de AG controle de armazenamento dos lipdeos de depsito e armazenamento dos lipdeos hepticos
6.1.1 CIDOS GRAXOS 6.1.1.1 BIOSSNTESE
Glicose convertida em piruvato, seguindo uma srie de reaes enzimticas, entrando, ento, na mitocndria onde participa do ciclo do cido ctrico . Na presena de grande quantidade de piruvato, o cido ctrico dentro da mitocndria convertido em acetil coenzima A , que a unidade bsica de 2C necessria para sntese de AG. via sinttica localizada primariamente dentro do hepatcito que o principal stio de lipognese de novo nas aves (em sunos principalmente o tecido adiposo). A sntese de AG requer a presena de H fornecido pelo NADPH e energia que suprida pelo ATP. Em aves o NADPH derivado da enzima mlica que cataliza a converso de cido mlico em pirvico. A via das pentoses est virtualmente ausente nesta espcie.
72
A sntese de AG a partir de carboidratos ou aminocidos, supe o catabolismo prvio destes at acetil CoA. O acetil CoA e malonil CoA condensam-se a um composto intermedirio, o butiril CoA. Este condensa-se juntamente com uma molcula de malonuil CoA para produzir caproil CoA.(figura 8) E, assim, essencialmente atravs da condensao dos AG com malonil CoA chega-se ao cido palmtico, que pode atuar como precursor de outras molculas saturadas. Tambm possvel a sntese de AG insaturados a partir do palmtico, a exceo do linolico e linolnico. Os AG assim formados unem-se ao glicerol, para formar os TG. 6.1.1.2 CATABOLISMO
A decomposio de um AG um processo oxidativo (figuras 9 e 10) . Os tomos de hidrognio so removidos pelas desidrogenases. Antes de iniciar a oxidao, que se processa na mitocndria, os AG so ativados por meio de esterificao com CoA. Depois da oxidao, dois tomos de carbono da extremidade carboxila so removidos na forma de acetil CoA. O resto da cadeia que ainda persiste em forma de ster de AG (acil CoA graxo) pode reiniciar todo o processo. O acetil CoA liberado durante a oxidao torna-se disponvel para ressntese de AG, esterides e cetonas ou pode participar do ciclo de Krebs. 6.1.2 TRIACILGLICERIS
Os TG so o grupo mais significativo de lipdeos do ponto de vista do metabolismo energtico dos animais. Eles podem ser fornecidos pela dieta ou sintetizados a partir de fontes no lipdicas (de novo), amplamente no fgado, tecido adiposo, glndula mamaria em lactao, nos rins, crebro e pulmes. 6.1.2.1 BIOSSNTESE
Duas vias gerais tm sido propostas para sntese de TG (figuras 11 e 12). A primeira envolve fosfotidato como intermedirio, enquanto a segunda no inclui intermedirios fosforilados. Em cada caso, os AG ativados (acil CoA graxo) so essenciais para formao de ligaes ster de gliceril. Em ambas as vias, um diacilglicerol (DG) produzido e, subseqentemente, esterificado para formar um TG. O -glicerol fosfato da via fosfatidato gerado pela reduo da diidroxiacetona fosfato (DHAP) da via glicoltica ou pela fosforilao com ATP pela glicerocinase. Tambm no fgado a DHAP pode ser esterificada com um acil CoA graxo antes da reduo do meio carbonil. A via fosfatidato predomina na sntese de TG em muitos tecidos, enquanto que o monoacilglicerol (MG) serve como substrato principal para sntese de TG na clulas epiteliais da mucosa durante a formao de quilomcrons (figura 13). 6.1.2.2 DEGRADAO
O catabolismo do TG (figura 14) a CO2 um importante gerador de energia utilizvel em animais. Os depsitos de TG so hidrolizados prontamente por lipases teciduais e os AG liberados so oxidados in situ ou transportados para outros tecidos sob a forma de complexos formados por AG-albumina. O fgado, o corao e os msculos esquelticos (figura 7) em repouso contam exclusivamente com a oxidao dos AG a CO2. O glicerol
73
liberado transportado, em primeiro lugar, para o fgado, formando-se -glicerol fosfato. Este metablito pode ser usado para sntese de acilglicerol ou oxidado a DHAP, para uso no metabolismo energtico ou na converso em glicose. 7 DIGESTO, ABSORO E UTILIZAO DE LIPDEOS 7.1 DIGESTO Uma caracterstica essencial dos lipdeos sua polaridade. Diz-se que uma molcula anfittica, quando possui simultaneamente um forte grupo hidroflico (carboxila) e outro hidrofbico (cadeia carbonada). Quando um lipdeo de alta polaridade pe-se em contato com a gua, tende a concentrar-se em pequenas associaes de molculas, denominadas micelas, orientando os grupos hidroflicos para o exterior e os hidrofbicos para o interior da micela. As caudas no polares da molcula (cadeia parafnica) esto dentro da micela, escondidas da gua, enquanto os grupos carboxila, negativamente carregados, esto expostos na superfcie da molcula e interagem com a fase aquosa. Tais micelas so um composto anfiptico, que permanecem uniformemente suspensos em gua, devido negatividade de suas cargas. Os lipdeos podem, ento, ser classificados segundo a sua polaridade, por sua maior ou menor capacidade para dissociar-se e formar micelas em meio aquoso: a) Polaridade muito baixa: tri- e diacilgliceris b) Polaridade baixa: cido graxo saturado de cadeia longa e esteris c) Polaridade mdia-alta: cidos graxos de cadeias curta e mdia; cidos graxos saturados de cadeia longa, monoacilgliceris e fosfolipdeos. A principal funo da digesto das gorduras converter, por hidrlise, os TG do alimento em compostos mais polares (monoacilgliceris e AG), potencialmente solveis no contedo digestivo. 7.1.1 DIGESTO PR-DUODENAL A digesto de gordura nos sunos parece iniciar-se no estmago mediante ao de uma esterase salivar e uma lipase gstrica (figura 15). Estas enzimas atuam seletivamente liberando cidos graxos de cadeia curta e mdia. Sua importncia quantitativa escassa em animais adultos e algo superior em leites, nos quais se observou que cerca de 26% dos TG so hidrolisados no estmago a MG e DG. As lipases salivar e gstrica possuem especificidade por ligaes com AG de cadeias mdia e curta. Tais AG podem ser absorvidos diretamente pela parede estomacal e isto apresenta grande importncia para o neonato como fonte imediata de energia. A gordura do leite contm maior proporo de AG de cadeias mdia e curta, na maioria das espcies.
74
A digesto pr-duodenal perde importncia com o avano da idade do animal. Em adultos, a medida que o HCl do estmago torna cido o contedo estomacal, a acidez paraliza a atividade das lipases gstrica e salivar, que atuam fundamentalmente em meio alcalino. A digesto gstrica da dieta importante porque a maioria dos constituintes dos lipdeos fazem parte dela. Tanto o pH baixo como a atividade proteoltica da pepsina atuam em conjunto para coalescer os lipdeos dos lipdeos. Os AG livres e os fosfolipdeos so parcialmente ionizados e atuam como anffilos em pH quase neutro, mas quando o pH do ambiente aquoso diminui para 1 a 2, eles ficam no dissociados e tornam-se hidrfobos, unindo-se com TG. Aminocidos hidrofbicos atuam como stios para agregao lipdica, quando vrios esto sequenciados em ligaes peptdicas. Porm, a pepsina hidrolisa estes stios, evitando ligaes nestas reas. A pepsina tambm hidrolisa protenas ligadas a lipdeos, tal como no germen dos gros. O efeito final da digesto no estmago (sunos) ou proventrculo-moela (aves) destruir a integridade estrutural do alimento e formar um pool de lipdeos. Falhas na digesto adequada de protenas da dieta podem resultar em absoro reduzida de AG, a partir do intestino delgado. Assim, valores de digestibilidade pela pepsina dos alimentos no so apenas ndices de disponibilidade de protenas, mas afetam, da mesma forma, a utilizao das gorduras. A coalescncia de lipdeos a um pH baixo no apenas une cada uma das classes, mas resulta em um ponto de fuso da mistura que a torna, geralmente, lquida a temperatura do corpo. Embora muitas gorduras animais sejam lquidas a temperatura corporal, a contribuio de gordura de origem vegetal e de baixos pontos de fuso, quando combinados, suficiente para resultar em reduo favorvel. 7.1.2 DIGESTO DUODENAL Os lipdeos so solubilizados na luz intestinal mediante a formao fsico-qumica de micelas lipdeo-sais biliares. Os sais biliares agem como detergentes, diminuindo a tenso superficial das solues aquosas, permitindo que molculas insolveis formem facilmente micelas compostas9 figura 16) . Estas micelas, tm a propriedade de solubilizar inmeras outras molculas de AG no polares, substncias afins aos lipdeos e vitaminas lipossolveis (AG, MG , DG e TG, colesterol, steres de colesterol, fosfolipdeos, vitaminas A, D, E e K). Os sais biliares acumulam-se na superfcie da gotcula de gordura, carregando negativamente a micela. A garga negativa atrai a colipase e a lipase pancretica. A funo da colipase fixar a lipase pancretica prxima a superfcie, a revelia dos cidos biliares que, ao contrrio, a deslocariam. Os sais biliares, a colipase e o clcio, que estimula a ao da lipase, formam um complexo por interao (micela mista). A lipase pancretica age especificamente sobre as ligaes steres primrias dos triacilgliceris, produzindo inicialmente 1,2 diacilglicerol e, finalmente, 2-monoacilglicerol. As micelas mistas so transportadas a superfcie das clulas epiteliais da mucosa, onde so absorvidas ao nvel de jejuno proximal. 7.2 ABSORO Os produtos derivados da hidrlise dos lipdeos so absorvidos, principalmente, pelas clulas da mucosa do jejuno proximal, ainda que se efetue uma absoro de menor
75
grau at a poro distal do intestino delgado. Esses produtos (AG de cadeia longa, colesterol livre, AG de cadeia curta, glicerol e monoacilgliceris) so incorpoprados ao citoplasma dos entercitos e rapoidamente utilizados na reestruturao dos triacilgliceris, steres de colesterol e fosfolipdeos (figura 15) . O glicerol e os AG de cadeia curta (com menos de 10C) se absorvem por meio de transporte passivo a corrente sangnea venosa mesentrica, de onde passam a corrente sangnea portal. Os demais lipdeos sofrem reesterificao dentro da clula para formar triacilgliceris que, juntamente com fosfolipdeos, colesterol e steres de colesterol so envolvidos por uma estrutura pseudo-membranosa de natureza protica o quilomcrom. Sob esta forma, os lipdeos abandonam a clula da mucosa por meio de uma pinocitose invertida, penetram nos quilferos e da so conduzidos ao sistema linftico que transporta os quilomcrons ao sangue. Atravs deste, os quilomcrons so transportados ao ducto torcico e, por fim, ao fgado, onde sero metabolizados. Nas aves o processo de reesterificao dos AG em triacilgliceris dentro da clula similar ao dos mamferos, entretanto a conduo dos lipdeos ao sistema portal o transporta ao fgado. Tanto nos mamferos quanto nas aves, os quilomcrons so captados pela fgado e tecido adiposo, onde so hidrolisados pela lipoproteina lipase at AG e glicerol. O fgado pode utiliz-los para fins catablicos ou anablicos, de modo similar aos nutrientes procedentes da digesto dos carboidratos. Em suma os AG e o glicerol podem: - incorporar-se ao tecido adiposo ou a glndula mamria, onde os AG so rapidamente esterificados a triacilgliceris; - incorpoprar-se ao tecido muscular onde podem armazenar-se temporariamente; - oxidar-se para sntese de ATP. As clulas do tecido adiposo hidrolizam os quilomcrons na sua superfcie exterior. Os AG e glicerol liberados formam TG no interior do lipcito. Ainda no interior das clulas adiposas, os TG so hidrolizados por lipases a AG e glicerol que, ao chegarem ao exterior da clula, so incorporados ao sangue. Assim, o tecido adiposo experimenta contnua renovao mediante este processo 7.3 UTILIZAO 7.3.1 DIGESTIBILIDADE A maior parte do contedo em lipdeos das fezes dos sunos, encontra-se na forma de sabes clcicos. Os sabes formam-se no intestino delgado preferencialmente com AG saturados de cadeia longa (16:0 e 18:0) e no so incorporados s micelas. O procedimento habitual de extrao dos lipdeos com ter no dilue os sabes, pelo que se requer um tratamento com cido prvio a extrao. A no utilizao deste mtodo at datas recentes h suposto uma superestimao da digestibilidade da gordura (sobretudo da mais saturada) nos primeiros trabalhos publicados sobre o tema.
76
A formao de sabes explica, da mesma forma, a observao de alguns autores de que um incremento do contedo de minerais da dieta afeta negativamente o coeficiente de digestibilidade da gordura (tabela 6) especialmente se esta saturada ou se o nvel de AG livres elevado.
Tabela 6. Efeito do contedo em minerais da dieta sobre a digestibilidade da gordura.(%) Nvel de minerais* 50 100 CD da gordura 75 71 CD c. Esterico 32 23 CD c. Lnolico 86 83 *expressos em % dos standards dinamarqueses de necessidades 150 69 19 81
Outro fator importante que determina a digestibilidade de uma gordura sua polaridade, isto , sua capacidade para formao de micelas. Por esta razo, o coeficiente de digestibilidade da gordura saturada inferior ao da insaturada ou a dos TG com AG de cadeias curta e mdia. Assim, quando se ingerem como AG livres, estima-se que a digestibilidade dos cidos caprico (10:0), palmtico (16:0) e esterico (18:0) ao redor de 100, 60 e 20% respectivemente.; a digestibilidade dos AG dos TG superior aos dos AG livres, devido polaridade aportada pelos monoacilgliceris que so gerados durante o processo de digesto. Analogamente, a digestibilidade do leo de pescado diminue ao aumentar o grau de hidrogenao (tabela 7) e das gorduras saturadas aumenta quando se administram misturas com gorduras mais digestveis. Tabela 7. Efeito do ponto de fuso sobre o coeficiente de digestibilidade aparente do leo de pescado (CDa , %) Ponto de fuso(C) CDa 32 72 38 72 44 61 50 53
O efeito do tipo de gordura sobre a digestibilidade mais pronunciado em leites que em sunos adultos. Os leites digerem facilmente o leite da porca, cujo contedo em gordura (altamente saturada, ver tabela 4) da ordem de 40%, porm tem dificuldades para utilizar a gordura saturada adicionada dieta pr-inicial (tabela 8); especula-se que a diferena est no maior grau de diviso das partculas gordurosas no leite e com maior freqncia de alimentao dos leites quando tomam leite que quando ingerem
77
rao. Como conseqncia, em dieta pr-incial recomenda-se utilizar gordura lctea, ou em sua falta, leos vegetais ou manteiga; o uso de olenas e sebo deve estar muito limitado em leites at 5 semanas e pode incrementar-se ao aumentar a idade do animal (tabela 8) 7.3.2 DEPOSIO DE GORDURA Durante os processos de absoro e transporte no se produzem alteraes na composio de AG, de modo que, em no ruminantes, existe grande semelhana entre a composio da gordura da dieta e a gordura corporal depositada a partir da anterior. No tecido adiposo sintetizam-se tambm triacilgliceris a partir de glicose procedente da digesto de carboidratos. A composio da gordura sintetizada por esta via caracterizase por elevado contedo de cidos palmtico, esterico e olico, que supe mais de 90% do total de cidos graxos. O aporte de gordura na dieta inibe a sntese de gordura a partir de hidratos de carbono. A composio final da gordura corporal ser, pois, uma mdia ponderada entre a gordura produzida de forma endgena a partir de glicose (sntese de novo) e a quantidade e composio dos AG da dieta. A adio de gordura dieta resulta em incremento nos depsitos de gordura corporal ou da gordura do leite. A adio de gorduras dieta de porcas eleva a concentrao energtica da rao e a produo de leite, isso reduz considervelmente a mortalidade de leites durante a lactao, especialmente dos que apresentam menor peso ao nascimento. A adio de gordura a dieta de frangos em crescimento aumenta a ingesto de alimentos e a produo. 7.3.3 MOBILIZAO DE RESERVAS Os TG armazenados como reserva de energia no tecido adiposo podem hidrolizar-se para serem utilizados por outros tecidos em funo das necessidades do organismo (figuras 17 e 18)). Assim, todos os TG corporais podem deslocar-se na forma de AG livres, at os msculos, ao oviduto e as glndulas mamrias nas fases de maior produo de leite, ou oxidar-se para produo de ATP nos animais subalimentados, ou aumentar a produo de calor em condies de baixas temperaturas. Por outro lado, o tecido adiposo metablicamente ativo, de modo que a gordura corporal no esttica, mas est continuamente mobilizando-se e ressintetizando-se. A vida mdia da gordura corporal no suno em fase de engorda estimada em 180 dias. Da, a possibilidade de alterar a composio da gordura depositada na carcaa, mediante manipulao do tipo de gordura adicionada dieta nas ltimas semanas de engorda. Os hormnios que regulam os processos de mobilizao da reserva de lipdeos (figura 19) atuam principalmente estimulando ou inibindo a atividade da enzima lipoprotena lipase nos diferentes tecidos animais. Com isso, altera seletivamente o fluxo de AG para outra funo metablica.
78
O metabolismo das gorduras apresenta tambm interaes importantes com algumas vitaminas: - a tiamina e a biotina intervm na sntese de gordura a partir de carboidratos - a riboflavina na oxidao dos AG - a colina requerida para biossntese de fosfolipdeos - o inositol um fator lipoprotico. 8 LITERATURA CONSULTADA BEORLEGUI, C.B., FERREIRA, W.M. Digestin y metabolismo de las grasas. ETSIA. Madri: 1990. 37p. CUNNINGHAM, J.G. Tratado de fisiologia Veterinria. Guanabara Koogan. 1993. 454p. DUKES, H.H., SWENSON, M.J. Fisiologia de los animales domesticos. 4 ed. Editora Aguilar. 1973. 1054p. vol.1.
METABOLISMO DIGESTIVO DAS PROTENAS E DO NITROGNIO NOS NORUMINANTES. INTRODUO. O termo protena foi proposto por Jns J. Berzelius no sculo XIX, a partir da raiz grega proteios, para descrever os componentes de primeira classe, ou de principal importncia que existem na natureza para a manuteno da vida. De certa maneira o significado da palavra ainda vlido porquanto as protenas exercem papis cruciais em, virtualmente, todos os processos biolgicos. I. FUNES. Catlise enzimtica. Nos sistemas biolgicos quase todas as reaes qumicas so catalisadas por macromolculas especficas chamadas enzimas, as quais exibem um enorme poder cataltico aumentando a velocidade de reao biolgica pelo menos um milho de vezes. Algumas destas reaes so muito simples (hidratao do CO2), entanto que outras, como a replicao dos cromossomos, so latamente complexas. O fato marcante quase todas as enzimas conhecidas so protenas; Transporte e armazenamento. Muitas molculas e ons pequenos so transportados por protenas especficas: a hemoglobina transporta oxignio nas hemcias, enquanto a mioglobina transporta a mesma molcula no msculo. O ferro transportado no plama sangneo pela transferrina e, armazenado no fgado ligado a uma outra protena, a ferritina; Movimento coordenado. A contrao muscular conseguida pelo movimento de deslizamento de dois tipos de filamentos proticos; os movimentos dos
79
cromossomos na mitose e a propulso de espermatozides por seus flagelos so produzidos por montagens contrteis constitudas de protenas; Sustentao mecnica. A alta fora de tenso da pele e do osso devida presena do colgeno, uma protena fibrosa; Proteo imunitria. Nos organismos vivos existem protenas altamente especficas chamadas de anticorpos que exercem um importante papel distinguindo entre o que e o que no prprio ao indivduo; Gerao e transmisso de impulsos nervosos. A resposta de clulas nervosas a estmulos especficos intermediada por protenas receptoras: a rodopsina, por exemplo, a protena receptora nos bastonentes na retina; nas sinapses existem protenas receptoras que podem ser acionadas por molculas pequenas altamente especficas como a acetilcolina participando assim da transmisso dos impulsos nervosos; Controle do crescimento, da diferenciao e a atividade das clulas. A expresso seqencial controlada da informao gentica essencial para o crescimento e a diferenciao ordenados das clulas. Nas bactrias, as protenas repressoras so elementos controladores importantes que silenciam segmentos especficos do DNA de uma clula. Nos organismos superiores, o crescimento e a diferenciao so controlados pelos fatores proticos de crescimento. Nos organismos muticelulares a atividade de diferentes clulas coordenada por hormnios como a insulina e o hormnio estimulante da tireide, que so protena. II. ESTRUTURA DAS PROTENAS. As protenas so definidas como compostos orgnicos complexos, colidais por natureza, de alto peso molecular e presentes em toda clula viva. Tal como as gorduras e carboidratos, as protenas contm carbono, hidrognio e oxignio. Alm disso, contm ampla e muito regular percentagem de nitrognio. A maioria delas contm enxofre alm de fsforo e ferro. A escala da composio elementar das protenas mais tpicas a seguinte: Carbono Oxignio Nitrognio Hidrognio Enxofre Fsforo Por cento 51,0-55,0 21,5-23,5 15,5-18,0 (mdia geral de 16,0%) 6,5- 7,3 0,5- 2,0 0,0- 1,5
Uma caracterstica marcante das protenas de todas as espcies, desde as bactrias at o homem, que estes componentes elementares das protenas encontram-se dispostos em estruturas tridimensionais bem definidas construdas a partir de um repertrio bsico de 20 aminocidos, que do a funo da protena. Isto significa que uma cadeia protica distendida ou disposta de maneira aleatria no tem atividade biolgica; ela conseqncia da sua conformao, ou seja, da disposio tridimensional de seus tomos na estrutura. III. PROPRIEDADES DAS PROTENAS.
80
As protenas podem combinar-se quimicamente com cidos e bases. Embora os grupos amino e carboxila das ligaes peptdicas no so funcionais nas reaes entre cidos e bases, todas as protenas contm grupos amino e carboxila livres, seja como unidades terminais ou nas cadeias laterais dos aminocidos. Consequentemente, as protenas, como os aminocidos, so substncias anfteras. Cada protena tem o seu ponto isoeltrico caracterstico, no qual so iguais as tendncias para as dissociaes cidas e bsicas e a protena precipitada por solues salinas o por lcool. Esta propriedade til para a separao e purificao das protenas. Todas as protenas em soluo tm propriedades coloidais. Variam no que concerne a sua solubilidade em gua, desde as queratinas, que so insolveis, at as albminas, que so solveis. Contudo, nenhuma protena solvel nos solventes graxos comuns, como o ter etlico e de petrleo. As protenas solveis podem ser precipitadas acrescentando soluo certos sais neutros (sulfato de sdio e de magnsio); caso que a soluo seja diluda, as protenas voltam a ser solveis. As protenas so compostos to lbeis que a presena de agentes fsicos, a aplicao de altas temperaturas e a silagem e a armazenagem prolongada podem alterar o seu estado natural (desnaturando-a) e, portanto, afetando o seu valor nutritivo. O exemplo mais conhecido de desnaturao(1) das protenas a coagulao, por calor, da clara de ovo. Embora a maioria das protenas so coaguladas pelo calor, so inmeros os agentes que produzem este fenmeno: cidos fortes, lcalis, lcoois, acetona, uria e sais de metais pesados.
(1).
A desnaturao toda alterao no-proetoltica da estrutura prpria de uma protena natural, que determina mudanas definidos nas propriedades qumicas, fsicas e biolgicas das protenas. Neste conceito no so includos os produtos prprios originados pela hidrlise natural das protenas. Os efeitos mais marcantes da desnaturao so observados nas mudanas das propriedades biolgicas; as enzimas, por exemplo, so inativas. As protenas, so especiais por serem capazes de reconhecer e interagir com molculas altamente diversas devido a sua alta capacidade na formao de superfcies e fendas complementares; o rico repertrio de 20 tipos de cadeias laterais nessas superfcies e fendas permite que as protenas formem pontes de hidrognio, ligaes eletrostticas e de van der Waals com outras molculas. Vejamos alguns exemplos desta capacidade: a mioglobina liga-se fortemente a um grupamento hemo quando sua cadeia polipeptdica est parcialmente enovelada; a aquisio desse grupamento permite hemoglobina ligar-se reversivelmente ao O2, que sua funo biolgica. As protenas tambm combinam-se com outras produzindo arranjos altamente ordenados como os filamentos contrteis no msculo. A ligao de molculas estranhas aos anticorpos, outros tipos de protenas, est no fundo da capacidade do sistema imunitrio de distinguir o que ou no prprio no organismo vivo. A expresso de muitos genes controlada pela ligao de protenas que reconhecem seqncias especficas de DNA. De um modo geral, o poder cataltico das protenas deve-se sua capacidade de ligar-se a molculas de substrato em orientaes precisas e de estabilizar estados de transio na produo e na quebra de ligaes qumicas. Finalmente, as mudanas de conformao transmitidas entre locais distantes nas molculas proticas so a base da capacidade das protenas de transferncia de energia e informao.
81
IV. CLASSIFICAO DAS PROTENAS. 1. SEGUNDO A SOLUBILIDADE. Segundo a forma, composio qumica e solubilidade as protenas so classificadas nos seguintes grupos: Protenas simples ou globulares. So protenas que produzem apenas aminocidos, ou seus derivados, quando solubilizadas em gua, cidos diludos, bases e lcool. Exemplos deste tipo so: Albuminas (albumina do ovo e do soro sangneo), histonas e protaminas: solveis em gua e coagulam-se ao se aquecer. Esto nos ovos, leite, sangue e inmeros vegetais; Globulina muscular e legumina do feijo e ervilhas: insolveis ou pouco solveis em gua, solveis em uma soluo neutra diluda de sais e coagulveis por aquecimento. Encontram-se nos ovos, leite e sangue; Glutelinas: solveis em cidos e bases diludos, sendo que as mais marcantes so as glutelinas do trigo; Prolamina: solvel em etanol ao 70%. O exemplo mais marcante deste grupo a zeina do milho. Protenas fibrosas. So protenas insolveis, muito resistentes s enzimas digestivas dos animais, formadas por cadeias filamentosas alongadas unidas por ligaes transversais. Deste grupo fazem parte: Colgenos: constituem 30% do total da protena do corpo animal; so as protenas mais importantes do tecido conectivo. Elastina: uma protena dos tecidos elsticos como tendes e artrias; Queratinas: so protenas muito ricas em cistina (aminocido com enxofre), presentes em plo, pele, unhas. Entanto que os colgenos e a queratina so insolveis em gua, a queratina, alm de ser altamente insolvel, uma protena indigestvel. Protenas conjugadas ou compostas. So protenas que, por hidrlise, liberam, alm dos aminocidos, grupos no-proticos chamados grupos prostticos. Cinco subgrupos so identificados neste tipo de protenas: Nucleoprotenas: so compostos de alto peso molecular constitudos por uma ou mais molculas proticas mais o cido nuclico como grupo prosttico. Esto presentes em germes de sementes e tecido glandular; Glicoprotenas (mucoprotenas): so protenas conjugadas com um ou vrios heteroglicanos como grupo prosttico. Na maioria dos casos os heteroglicanos contm glucosamina, galactosamina, galactose e manose. Formam parte das secrees das mucosas atuando como lubrificantes em inmeros locais do organismo;
82
Lipoprotenas: trata-se de uma protena simples que contm lpides (lecitina, colesterol) no seu radical. So os componentes principais das membranas celulares realizando uma funo bsica no transporte de lpides; Fosfoprotenas: protenas que formam complexos com um radical fosfrico diferente a um cido nuclico ou a um fosfolpide. Os exemplos deste grupo so: a casena do leite e a fosfovitina da gema do ovo; Cromoprotenas: compostos da molcula protica com um pigmento como grupo prosttico. A hemoglobina, hemocianina, citocromo e flavoprotenas so exemplos deste grupo. No caso da hemoglobina, a protena globina est combinada com um composto que contm ferro, o grupo HEM ou hematina. Protenas derivadas. Consistem de compostos gerados a partir da quebra e alterao de grupos de protenas conjugadas, produzida pela ao do calor, enzimas ou agentes qumicos. Constitui um grupo grande subdividido em menores representados pelos diferentes graus de descomposio, isto : derivados protenicos primrios, proteanos, metaprotenas, protenas coaguladas, derivados protenicos secundrios, proteoses, peptonas e peptdios. As protenas hidrosolveis enovelam-se em estruturas compactas com interior apolar. Em vrias protenas, como a mioglobina por exemplo, o enovelamento da cadeia principal complexo e sem simetria. Contudo, em todas elas existe um princpio unificador surgido da distribuio das cadeias laterais. O fato marcante nestas protenas que o interior est constitudo quase por completo de radicais apolares (Leu, Val; Met, e Fen), sendo que esto ausentes os radicais polares (Asp, Glu, Lis e Arg). O exterior da protena, pelo contrrio, contm radicais tanto polares quanto apolares. Essa distribuio contrastante de radicais polares e apolares revela uma faceta essencial da arquitetura protica. Em um meio aquoso, o enovelamento da protena comandado pela forte tendncia dos radicais hidrfobos em ser excludos da gua; importante salientar que a gua altamente coesiva e que os grupamentos hidrfobos so termodinamicamente mais estveis quando aglomerados no interior da molcula do que quando estendidos no ambiente aquoso. Por isso a cadeia polipeptdica dobra-se espontaneamente, para que suas cadeias laterais hidrfobas fiquem soterradas entanto que na superfcie fiquem suas cadeias polares carregadas. As protenas que integram as membranas biolgicas so planejadas de maneira diferente das que so solveis em meios aquosos. A barreira de permeabilidade das membranas formada de lipdeos, que so altamente hidrfobos; por isso, a parte de uma protena que atravessa essa regio deve ter um exterior hidrfobo. 2. SEGUNDO A FUNO BIOLGICA. Enzimas: pepsina, tripsina, ribonuclease; Protenas transportadoras: hemoglobina, albumina de soro, mioglobina 1lipoprotena; Protenas nutritivas e de reserva: gliadina (trigo), ovoalbumina (clara do ovo), casena (leite), ferritina; Protenas contrteis ou de movimento: actina, miosina, tubulina e dinena; Protenas estruturais: queratina, fibrona, colgeno, elastina e proteoglicanas; Protenas de defesa: anticorpos, fibrinognio, trombina, toxina, botulnica e diftrica, veneno de serpentes e ricina;
83
Protenas reguladoras: repressores.
insulina,
hormnio
do crescimento, corticotropina e
V. COMPOSTOS NITROGENADOS NO PROTICOS. A maior parte do material estrutural das clulas que contm nitrognio de origem protica. Contudo, as clulas animais e vegetais tm quantidades variveis de compostos nitrogenados que no fazem parte das protenas. Nas anlises de laboratrio esses compostos tm sido classificados, de forma geral, como compostos nitrogenados no proticos, sendo que constituem este grupo: alantonina, amidas (asparagina, glutamina, uria), aminas (histaminas), aminocidos livres, amnia, colina, creatina, creatinina, nitritos, nitratos, oxitocina, peptdeos, pigmentos, purinas, pirimidinas, cido rico, glutatione, vitaminas, alcalodes. VI. NVEIS DE ESTRUTURA NA ARQUITETURA DAS PROTENAS. Na estrutura das protenas citam-se freqentemente quatro nveis: Estrutura primria. a seqncia de aminocidos e a localizao de dissulfetos, se houver ao longo das cadeias de polipeptdeos das protenas. A estrutura primria , portanto, a descrio completa das conexes covalentes de uma protena. Estrutura secundaria. Refere-se conformao da cadeia de aminocidos gerada ao se formarem as pontes de hidrognio entre os grupos NH e carbonila dos aminocidos que esto perto uns dos outros na seqncia linear. Algumas dessas relaes estricas so de tipo regular, originando uma estrutura peridica encontrada sob a forma de -hlice ou de folha pregueada , ou pode ser irregular e, para tanto, encontra-se enovelada ao acaso. Estrutura terciria. Refere-se ao arranjo espacial de aminocidos que esto bem longe na seqncia linear. difcil estabelecer a linha divisria entre a estrutura secundria e terciria. As protenas que contm mais de uma cadeia polipetdica exibem mais um nvel de organizao estrutural, no qual cada cadeia chamada de subunidade. A interao entre os grupos R dos aminocidos nas cadeias da estrutura secundria determinam folhas e dobras na cadeia de polipeptdeos, proporcionando a cada protena sua atividade biolgica. Estrutura quaternria. As protenas possuem estrutura quaternria se contm mais de uma cadeia de polipeptdeos. As foras que estabilizam estas estruturas so pontes de hidrognio e ligaes eletrostticas formadas entre as molculas das superfcies das cadeias de polipetdeos. Estudos recentes de conformao, funo e evoluo das protenas revelaram a presena de mais dois nveis de organizao: Estrutura supersecundria: refere-se a aglomerados de estrutura secundria. Em muitas protenas, por exemplo, encontrada como uma fita separada de outra fita por uma -hlice;
84
Domnios: so unidades globulares compactas constitudas por 100 a 400 radicais de aminocidos nas que as cadeias polipeptdicas esto enoveladas em duas ou mais regies que podem estar unidades por um segmento flexvel de cadeia peptdica. VII. AS CADEIAS POLIPEPTDICAS ESTO FORMADAS POR AMINOCIDOS UNIDOS POR LIGAES PEPTDICAS. As protenas tm uma seqncia de aminocidos definida geneticamente, sendo que o grupamento -carboxila de um aminocido unido ao radical -amino de outro aminocido por uma ligao petdica ou ligao amida. Cada um dos 20 aminocidos das protenas codificado por uma ou mais seqncias especficas de trs nucleotdeos. As protenas de todos os organismos vivos, por sua vez, so sintetizadas a partir de seus aminocidos constituintes utilizando o mesmo mecanismo. A formao de um dipeptdeo a partir de dois aminocidos gera a perda de uma molcula de gua. O equilbrio dessa reao para a o lado da hidrlise (termodinamicamente favorvel), em vez da sntese (requer a entrada de energia livre). Muitos aminocidos so unidos por ligaes peptdicas formando uma cadeia polipetdica, no ramificada. Em um polipeptdeo uma unidade de aminocido pode ser chamada de radical ou resduo. Uma cadeia polipetdica tem direo, porque suas unidades constitutivas, os grupamentos -amino e -carboxila, tm extremidades diferentes. Por conveno internacional, a cadeia se inicia com a extremidade amnica, e por isso a seqncia de aminocidos da cadeia escrita comeando com o radical amino-terminal; desse modo, por exemplo, no tripetdeo Ala-Gli-Trp, a alanina o amino-terminal e o triptofano o carboxi-terminal. Uma cadeia polipeptdica constituda de uma parte repetida regularmente, chamada de cadeia principal, esqueleto ou espinha dorsal, e de uma parte varivel compreendo as diferentes cadeias laterais. Na natureza a maioria das cadeias polipeptdicas contm entre 50 e 2000 aminocidos. Se o peso molecular mdio de um radical de aminocido em torno de 110, o peso molecular de grande parte das cadeias polipetdicas est entre 5500 e 220000 (2). Algumas protenas contm pontes dissulfeto. Essas interligaes entre as cadeias ou entre as partes de uma cadeia so formadas pela oxidao de radicais de cistena. O dissulfato gerado chamado de cistina. As protena intracelulares geralmente no tm pontes dissulfeto, ao passo que as extracelulares freqentemente contm vrias. Em algumas protenas, as fibras de colgeno no tecido conjuntivo e os cogulos de fibrina por exemplo, tambm esto presentes interligaes no-sulfuradas derivadas das cadeias laterais da lisina. O conjunto bsico dos 20 aminocidos pode ser modificado aps a sntese de uma cadeia polipeptca o que traz aumento das suas capacidades; por exemplo, os terminais amnicos de muitas protenas so acetilados, tornando-as mais resistentes degradao. Existem inmeros outros exemplos que mostram que a modificao e a clivagem das protenas conferem novas capacidades. No colgeno recm sintetizado muitos radicais de prolina so hidroxilados formando hidroxiprolina; a adio destes radicais estabilizam a fibra do colgeno. Os anticorpos, que tambm so protenas, adquirem cadeias
85
glicdicas em determinados radicais de asparagina; fosfosserina e fosfotreonina so os mais comuns aminocidos modificados em protenas. Muitos hormnios, como a adrenalina, alteram a atividade de enzimas estimulando a fosforilao dos aminocidos hidroxilados serina e treonina. Fatores do crescimento, como a insulina, agem disparando a fosforilao do radical hidroxila da tirosina, formando fosfotirosina. Muitas outras protenas so clivadas e aparadas aps a sntese. Algumas enzimas digestivas, por exemplo, so sintetizadas como precursores inativos que podem ser armazenados com segurana no pncreas. Aps serem liberadas no intestino, esses precursores so ativados por clivagem das ligaes peptdicas. Na coagulao sangnea, o fibrinognio solvel transformado em fibrina insolvel por clivagem das ligaes peptdicas. Vrios hormnios peptdicos, incluindo o adrenocorticotrpico, vm da ciso de uma s protena precursora grande.
(2).
A massa de uma protena tambm pode ser expressa em unidades dlton, sendo que um dlton igual a uma unidade de massa muito prxima da do tomo de hidrognio (precisamente igual a 1,0000 na escala de massa atmica). Uma protena com um peso molecular de 50.000 tem uma massa de 50.000 dltons, ou 50 kd (quilodltons). Um kd (quilodlton) uma unidade de massa igual a 1.000 dltons. A unidade dlton foi denominada assim em homenagem a John Dalton que desenvolveu a teoria atmica da matria.
A unidade peptdica rgida e plana. O hidrognio do grupamento amina substitudo quase sempre trans (oposto) em relao ao oxignio do grupamento carbonila. No h liberdade de rotao em torno da ligao entre o tomo de carbono da carbonila e o de nitrognio da unidade peptdica porque essa ligao tem um carter parcial de ligao dupla. Em contrate, a ligao entre o tomo de carbono e o tomo de carbono da carbonila uma ligao simples pura. A unio entre o tomo de carbono e o tomo de nitrognio pepetdico tambm uma ligao simples pura. Consequentemente, h um grande grau de liberdade de rotao em torno dessas ligaes de cada lado da unidade pepetdica rgida. Nas cadeias polipeptdicas existem trs conformaes com repetio regular chamadas de -hlice, de folha pregueada e a hlice do colgeno. A -hlice uma estrutura em basto. A cadeia principal polipeptdica densamente enrolada forma a parte interna do basto, e as cadeias laterais estendem-se para afora em um arranjo helicoidal. A hlice estabilizada quando o grupamento CO de cada aminocido faz ponte de hidrognio com o NH do aminocido que est situado quatro radicais frente na mesma cadeia polipeptdica. A folha pregueada difere muito da -hlice porque uma folha em vez de um basto, quase toda estendida, em vez de fortemente enrolada, e estabilizada por pontes de hidrognio entre os grupamentos NH e CO em cadeia polipetdicas diferentes. Na folha pregueada as cadeias adjacentes podem correr na mesma direo (folha paralela) ou em direes opostas (folha antiparalela). As cadeias polipeptdicas podem mudar de direo fazendo voltas . Em sua maioria, as protenas tm formas compactas, globulares, devidas a numerosas reverses da
86
direo de suas cadeias polipeptdicas. As anlises das estruturas tridimensionais de numerosas protenas revelaram que muitas dessas reverses so produzidas porque o grupamento CO do aminocido n de um polipeptdeo faz ponte de hidrognio com o grupamento NH do aminocido (n+3). Desse modo, freqentemente voltas conectam fitas antiparalelas e cadeias polipeptdicas podem reverte abruptamente sua direo. VIII. AMINOCIDOS. Os aminocidos so os produtos gerados ao se hidrolisar as protenas mediante enzimas, cidos ou lcalis. Ainda que existem na natureza mais de duzentos aminocidos, somente encontram-se entre 18 e 22 tipos nas protenas animais e desses aproximadamente 12 e 14 so necessrios nas dietas destinadas para os sunos e aves, respectivamente. Os aminocidos variam em tamanho, forma, carga, capacidade de formao de pontes de hidrognio e reatividade qumica. A notvel gama de funes exercidas pelas protenas originada pela diversidade e a versatilidade desses 20 tipos de blocos de construo. Um -aminocido um cido orgnico formado de um grupamento nitrogenado bsico, que geralmente uma amina (-NH2), um grupamento carboxila (-COOH), um tomo de hidrognio e uma cadeia lateral ou radical, R, ligados a um tomo de carbono, chamado por ser o adjacente ao grupamento carboxila (Fig. 1). O radical que se liga ao carbono pode ser desde um simples tomo de hidrognio, como na glicina, at compostos aromticos ou alifticos. 1. PROPRIEDADES DOS AMINOCIDOS. A presena de um grupamento amino e um outro carboxilo faz que os aminocidos tenham caracter anftero, ou seja, possuem propriedades cidas e bsicas. As molculas que apresentam este tipo de caracterstica, podem se encontrar como molculas sem carga, como ons bipolares com cargas inicas contrrias ou em misturas com ambos estes tipos. Em soluo de pH neutro os aminocidos esto predominantemente na forma de ons dipolares ou zwitterions (3), em vez de molculas na-ionizadas. Na forma dipolar, o grupamento amino est protonado (NH3+) e a carboxila dissociada (-COO) (Fig. 1).
(3)
. Zwitter uma palavra que vem do alemo e significa hermafrodita. Diz-se de o ser que possui rgos reprodutores dos dois sexos.
NH3+ l H-C-COO l R Estrutura bsica do aminocido
NH2 l H-C-COOH l R
NH2 l H-C-COO l R Forma em on
Forma no-ionizada dipolar ou zwitteron
Figura 1. Estrutura das formas no-ionizadas e on dipolar de um -aminocido.
87
O estado de ionizao de um aminocido varia com o pH (Fig 2). Em soluo cida (pH de 1, por exemplo), a carboxila est na forma no-ionizada (COOH) e a amina est ionizada (-NH3+). Em soluo alcalina (pH de 11), a carboxila est ionizada (COO) entanto que a amina esta na sua forma no-ionizada (NH2). Para cada aminocido existe um valor de pH no qual eletricamente neutro; esse valor chamado de ponto isoeltrico. Para a glicina, por exemplo, o ponto mdio da primeira ionizao acontece quando o pH 2,3 e da amina 9,6; isto significa que na glicina o pk da carboxila 2,3 e o da amina 9,6. Sete dos 20 aminocidos tm cadeias laterais facilmente ionizveis. NH3+ l H-C-COOH l R H
+
NH3+ l H-C- COO l R
H+
NH2 l H-C-COO l R Forma predominante em pH 11
Forma predominante Forma predominante em pH 1 em pH 7
Figura 2. Estados de ionizao de um -aminocido em funo do pH. Por apresentarem os aminocidos um caracter anftero, atuam como tampes, resistindo s mudanas de pH. O arranjo tetradrico dos quatro grupamentos diferentes em torno do carbono confere atividade tica aos aminocidos. As duas imagens especulares so chamadas de ismeros L e D, sendo que s os aminocidos L so constituintes das protenas, e as enzimas proteolticas animais atacam apenas as ligaes que apresentam esta mesma configurao. A configurao determinada em relao com o D-gliceraldedo, utilizado como substncia padro. Dos 20 aminocidos a glicina especial por ser opticamente inativa. Para facilitar uma comunicao concisa, os aminocidos so freqentemente designados por uma abreviao de trs letras, ou por um smbolo com uma letra. As abreviaes dos aminocidos so as trs primeiras letras de seus nomes, exceto para o triptofano (Trp), asparagina (Asn), glutamina (Gln) e isoleucina (Ile). Os smbolos para os aminocidos menores so as primeiras letras de seus nomes (G, para a glicina; L, para a leucina) (Tab 1). 2. CLASSIFICAO DOS AMINOCIDOS. 2.1. AMINOCIDOS ALIFTICOS. O mais simples dos aminocidos a glicina, que s tem um tomo de hidrognio como cadeia lateral. Logo vem a alanina, que apresenta um radical metila (-NH3+) como cadeia lateral. Existem cadeias laterais hidrocarbonadas maiores, com trs e quatro tomos de carbono, nos aminocidos valina, leucina, isoleucina e prolina. Essas cadeias laterais alifticas maiores so hidrfoba, ou seja, tm averso gua e tendem a se aglomerar, formando estruturas compactas com poucos orifcios.
88
TABELA 1. ABREVIAES UTILIZADAS PARA DESIGNAR OS AMINOCIDOS. Aminocido Abreviao com trs letras Ala Arg Asn Asp Asx Cys Fen Gly Glu Gln Glx His Ile Leu Lys Met Pro Ser Thr Tre Trp Val Smbolo de uma letra A R N D B C F G E Q Z H I L K M P S Y T W V
Alanina Arginina Asparagina Aspartato Aspartato ou Asparagina Cistena Fenilalanina Glicina Glutamato Glutamina Glutamato ou Glutamina Histidina Isoleucina Leucina Lisina Metionina Prolina Serina Tirosina Treonina Triptofano Valina
A prolina tambm tem uma cadeia lateral aliftica, porm diferencia-se dos outros aminocidos por ter sua cadeia lateral ligada tanto ao carbono quanto ao nitrognio; a estrutura originada deste arranjo influencia profundamente a arquitetura das protenas. A prolina contm uma amina secundria em vez da primria, o que torna-a um iminocido, que no se nega a ficar exposto gua. 2.2. AMINOCIDOS COM CADEIAS LATERAIS AROMTICAS. Trs aminocidos fazem arte deste grupo: a fenilalanina, o triptofano e a tirosina. A fenilalanina, contem um anel fenil ligado a um radical metileno (-CH2-). O triptofano tem um anel indlico ligado ao radical metileno , sendo que essa cadeia lateral contm, alm dos tomos de carbono e hidrognio, um tomo de nitrognio. A fenilalanina e o triptofano so altamente hidrfobos. O anel aromtico da tirosina contm um radical hidroxila, o que torna a tirosina menos hidrfoba que a fenilalanina. Os anis aromticos destes trs aminocidos contm nuvens deslocadas de eltrons que permitem sua interao com outros sistemas e a transferncia de eltrons.
89
2.3. AMINOCIDOS COM ENXOFRE NAS CADEIAS LATERAIS. Existem dois aminocidos que apresentam enxofre nas suas cadeias laterais: a cistena e a metionina. Entanto a cadeia lateral da cistena contm um radical sulfidrila (-SH) altamente reativo, a metionina apresenta um tomo de enxofre em uma ligao tio-ter (-S-CH3). Em ambos os aminocidos as cadeias laterais sulfuradas so hidrofbas. 2.4. AMINOCIDOS COM RADICAIS HIDROXILA NAS CADEIAS LATERAIS. Dois aminocidos, a serina e a treonina, contm radicais hidroxila nas suas cadeias laterais, o que tornam-nas mais hidrfilas. A serina e a treonina so tidas como verses hidroxiladas da alanina e a valina, respectivamente. A treonina, como a isoleucina, contm dois centros de assimetria; todos os outros aminocidos, exceto a glicina, contm um s centro assimtrico, o carbono . A glicina especial por ser opticamente inativa. 2.5. AMINOCIDOS COM CADEIAS LATERAIS MUITO POLARES (AMINOCIDOS BSICOS). Em pH neutro, a lisina e a arginina, tm carga positiva, o que os torna altamente hidrfilos. Entre os 20 aminocidos estes dois so os que tm as cadeias laterais mais longas . A histidina, o outro aminocido que faz parte do grupo, pode estar ou no com carga positiva, dependendo do ambiente do local; de fato, a histidina freqentemente encontrada nos centros ativos das enzimas, onde o anel imidazlico pode variar entre esses estados catalisando a formao e a ruptura de ligaes. 2.6. AMINOCIDOS COM CADEIAS LATERAIS CIDAS. Nos 20 aminocidos, existem dois, o cido asprtico e o cido glutmico, que contm cadeias laterais cidas. Na realidade esses dois aminocidos so geralmente chamados de aspartato e glutamato para salientar que, sob condies de pH fisiolgico, suas cadeias laterais esto quase sempre com carga negativa. A glutamina (o termo mais conveniente seria glutamida visto que trata-se de um amida do glutamato e no de uma amina) e a asparagina, que contm um grupamento amida terminal em vez de carboxila, so derivados no-carregados do glutamato e do aspartato. Alm da classificao qumica os aminocidos tambm podem ser agrupados seguindo critrios metablicos e nutricionais. Quanto ao metabolismo animal os aminocidos podem ser classificados em glucognicos, glucocetognicos e cetognicos. Dependo de critrios nutricionais os aminocidos podem ser tidos como limitantes, esssenciais e no essenciais. Alm dos aminocidos j registrados, existem alguns outros no proticos que participam do metabolismo animal, mas no participam de molculas proticas: Citrulina e a ornitina, por exemplo, participam no ciclo da uria na via metablica da sntese da ornitina; Betalanina, um ismero da alanina, faz parte do cido pantotnico e, portanto, da coenzima A e da protena carreadora de acila; Creatina, uma amina quaternria derivada da glicina, faz parte do fosfato de creatina que participa no armazenamento da energia.
90
IX. METABOLISMO DIGESTIVO DAS PROTENAS. As protenas, nas suas condies naturais, apresentam uma estrutura tridimensional que oferece poucas ligaes susceptveis ao ataque das enzimas proteolticas produzidas pelo prprio animal, alm de estarem em uma tal forma que no apropriada para atravessar as membranas da mucosa do trato gastrointestinal; consequentemente, precisam se submeter a um processo que vise que seus constituintes mais simples, os aminocidos, possam ser liberados e absorvidos. Este processo, conhecido como digesto, inclui atividades qumicas e enzimticas, alm da prpria absoro dos aminocidos. As enzimas associadas ao processo digestivo das protenas nos no-ruminantes so registradas na Tabela 2. 1. DIGESTO NO ESTMAGO. Nos no-ruminantes o processo de digesto das protenas comea no estmago do animal com a secreo do suco gstrico, composto principalmente por gua, pepsinognio, sais inorgnicos, mucina, cido clordrico e o fator intrnseco, importante para a absoro da vitamina B12. O contedo de cido clordrico do suco gstrico varia como conseqncia de inmeros fatores, porm est em torno de 0,1 N, valor suficiente para abaixar o pH do estmago at 2,0.
TABELA 2. RGOS E ENZIMAS QUE INTERVM NA DIGESTO DAS PROTENAS NOS NO-RUMINANTES. RGO ESTMAGO ENZIMA (1) Pepsina Renina (Quimosina) Tripsina PNCREAS SUBSTRATO Endopeptdeos de aminocidos aromticos Casena PRODUTOS FINAIS Proteoses e peptdeos Paracaseinato de Ca Polipeptdeos Peptdeos Peptdeos Aminocidos livres
Endopeptdeos de arginina e lisina Quimotripsina Endopeptdeos de fenilalanina, tirosina e metionina Peptidase pancretica Ligaes peptdicos com aminocidos neutros Carboxipeptidases A Exopeptdeos de fenilalanina, tirosina, triptofano, leucina
91
Carboxipeptidases B MUCOSA INTESTINAL Di e Tripeptidases Aminopeptidases Carboxipeptidases Polinucleotidases (Desoxirribonuclease, ribonuclease) Nucleosidases Fosfatases
Exopeptdeos de aminocidos bsicos Di e tripeptdeos Peptdeos com grupos -NH2 Terminal Peptdeos com grupos -COOH terminal cidos nuclicos (ADN, RNA) Ligao entre a ribose e a base nitrogenada
Aminocidos livres Aminocidos livres Aminocidos livres Aminocidos livres Nucletide Purinas e pirimidinas
Liberao do H3PO4 da ribose ou da desoxirribose (1). Algumas das enzimas digestivas so secretadas como precursores inativos sendo ativados sob a influncia de outras enzimas, pH ou diante a presena de ons. A ativao destes precursores tal vez requer o desmembramento de uma seo peptdica que, provavelmente, obtura o centro ativo da enzima; assim,por exemplo, entanto o pepsinognio tem peso molecular de 42000, na pepsina de 34000. O cido clordrico desmembra uma seo peptdica de baixo peso molecular do pepsinognio, que a forma inativa da pepsina, e com isto ativado na forma de pepsina. Nos sunos tm sido achados quatro tipos de pepsina, sendo que so ativas quando o pH estomacal atinge valores entre 2,0 e 3,5. A pepsina se ativa, atacando com preferncia as ligaes peptdicas adjacentes aos aminocidos aromticos fenilalanina, triptofano e tirosina; contudo, a pepsina tem forte atividade sobre as ligaes que envolvem aminocidos dicarboxlicos (cido glutmico e cido asprtico). Embora nos mamferos jovens a protena do leite degradada pela renina (quimosina), a pepsina tambm exerce uma potente ao coagulante sobre este nutriente. No estmago, os produtos da digesto das protenas so fundamentalmente protenas parcialmente digeridas e polipeptdeos de cadeias com comprimento varivel. Nas aves, limitada a digesto das protenas do alimento no ventrculo ou estmago glandular visto que a ingesta atravessa muito rapidamente por esse local do trato digestivo (14 segundos). Embora a moela no secreta enzimas digestivas, a ao da pepsina continua a agir durante a passagem do alimento ao ventrculo, moela ou estmago glandular pelo tempo que a ingesta ali permanece. Anatomicamente o coelho tem um estmago e ceco bastante desenvolvidos (com capacidade de conter cerca de 80% da digesta) e esto bem adaptados fermentao microbiana. No coelho adulto o estmago apresenta uma capacidade mdia de 500ml que considervel ao se comparar com o de outros herbvoros no-ruminates como o eqino; alm de mais, normalmente no se encontra vazio e tem a particularidade de ser pouco contrtil. Estas particularidades permitem aos alimentos e s fezes moles
92
permanecerem o tempo suficiente para manter o pH favorvel ao metabolismo bacteriano. As fezes cecotrofas so consumidas diretamente do nus e ingeridas ainda serem mastigadas. No estmago estas no so misturadas imediatamente com o contedo gstrico, sendo que permanecem intatas na regio do fundus entre 6 e 8 horas. Durante este perodo as fezes resistem as aes mecnicas, qumicas e enzimticas graas s camadas mucosas que as cobrem, convertendo-se em pequenas focos de fermentao. A presena de um sistema tampo-fosfato no interior das fezes garantem um pH favorvel (4-6) para o metabolismo das bactrias apesar de que o resto do contedo estomacal seja cido (pH 1-1,5). A atividade microbiana termina quando a mucosa protetora que cobre as cecotrofas so destrudas. As pesquisas tm mostrado que no antrum do estmago secretada uma substncia bacterioltica ativada pela pepsina permitindo, em conseqncia, que as enzimas proteolticas ataquem as protenas contidas no estmago. O eqino um herbvoro no-ruminante. Como no coelho, o eqino tem um trato gastrointestinal caracterizado pela presena de um intestino grosso bastante desenvolvido, mas, que a diferena deste apresenta um estmago com uma limitada capacidade para a armazenagem de alimento (15-18l) o que, portanto, obriga ao seu freqente esvaziamento (6-8 vezes em um dia) e rpida passagem do alimento por este local, limitando o alcance das aes mecnicas, qumicas e enzimticas sobre a digesto das protenas, porm sem limitar uma certa atividade microbiana fermentativa.
2. DIGESTO DAS PROTENAS NO INTESTINO DELGADO. No intestino delgado dos no-ruminantes so semelhantes os processos digestivos sobre as protenas do alimento; consequentemente, o modelo proposto a seguir vlido para descrever estes processos nas aves, coelhos, eqinos e sunos. As protenas parcialmente digeridas e os polipeptdeos gerados pela digesto estomacal das protenas passam ao intestino delgado e misturam-se com as secrees do duodeno, fgado e pncreas. Ao contrrio da pepsina, as enzimas proteolticas que agem no intestino delgado tm um pH timo para serem ativas de 7 a 9. O zimgeno inativo ou tripsinognio, ativado pela ao de uma enteroquinase produzida pela mucosa intestinal; assim que ativado, o tripsinognio transformado em tripsina, que por sua vez, e por um mecanismo autocataltico, tambm ativa o zimgeno. Parece que este processo de ativao da enzima se produz quando no zimgenio liberado um hexapeptdeo do grupo terminal do tripsinognio. A tripsina altamente especfica e por isso s age sobre as ligaes peptdicas onde esto presentes a lisina e a arginina; alm disso, a tripsina tambm participa nos processos que visam a ativao do quimotripsinognio em quimotripsina. Esta nova enzima, tambm altamente especfica, ataca as ligaes peptdicas nas que intervm os grupos carboxilo da tirosina, triptofano, fenilalanina e leucina. Novamente a tripsina participa na ativao de outro grupo de enzimas, as procarboxipeptidases, transformando-as em enzimas proteolticas carboxipeptidases. Este novo grupo de enzimas age no intestino delgado atacando os peptdeos da parte final da cadeia, escindindo os aminocidos terminais que possuem um -carboxilo livre. Por esta razo as carboxipeptidases so enzimas tidas como exopeptidases, entanto que a tripsina e a
93
quimotripsina so endopetidases, o que significa que sua atividade est orientada a agir sobre as ligaes peptdicas que ficam na parte interna do polipeptdeo. Os cidos nuclicos, o DNA e o RNA so hidrolisados pelas polinucleotidases, a desoxirribonuclease e a ribonuclease, respectivamente. Estas enzimas catalisam o rompimento das ligaes ster entre a ribose e o cido fosfrico dos cidos nuclicos, sendo que os produtos finais desta atividade so os nucleotdeos. Por sua parte, as nucleosidases atacam as ligaes entre a ribose e as bases nitrogenadas, liberando as purinas e as pirimidinas. As fosfatases completam a hidrlise, liberando o cido ortofosfrico da ribosa e da desoxirribose. Finalmente, a hidrlise dos peptdeos pequenos at a libertao dos aminocidos realizada pelas enzimas secretadas junto s clulas desprendidas da mucosa intestinal. Grande parte dos peptdeos pequenos so hidrolisados por aminopeptidases, especificamente na ligao peptdica adjacente ao grupo amino livre. Embora grande parte desta hidrlise acontece na superfcie externa da clulas epiteliais, tambm se apresenta a absoro celular de alguns peptdeos e sua posterior degradao pelas enzimas presentes no citoplasma. Os dipeptdeos, por sua parte, so degradados pelas dipeptidases at aminocidos. Nos eqinos embora as aes digestivas sejam de curta durao no intestino delgado, esta regio representa um segmento importante do trato gastrointestinal para a digesto das protenas, mais ainda se levamos em considerao que sua capacidade quatro vezes superior do estmago. 3. TRANSPORTE DOS PRODUTOS DOS PROCESSOS DIGESTIVOS DAS PROTENAS DO LUME MUCOSA INTESTINAL. Sabe-se que nos mamferos a absoro por pinocitose de protenas intactas ocorre at poucas horas aps do nascimento. As protenas assim absorvidas so vitais para as crianas, bezerros, leites e cordeiros visto que desta maneira recebem os anticorpos que necessitam e que no foram passados pela placenta materna. A maioria dos neonatos perde rapidamente esta capacidade, porm tambm se admite que em alguns casos continua durante algum tempo. Nos animais adultos, consequentemente, no h absoro de protenas intactas; s possvel a absoro no intestino delgado de aminocidos livres, peptdeos e oligopeptdeos (constitudos entre dois e seis aminocidos). Nos animais adultos, por enquanto, tm sido elucidado que existem quatro sistemas diferentes de transporte ativo de aminocidos do lume mucosa intestinal: Para os aminocidos neutros Gly, Ala, Ser, Thr, Vla, Leu e Ile; Para os aminocidos bsicos Arg, Lys, Cys e His; Para os aminocidos cidos cido asprtico (asparagina) e cido glutmico (glutamina); Para a Gly e os iminocidos. Sabe-se que na presena de tais sistemas de absoro de aminocidos est a explicao da competio existente entre os aminocidos de estrutura semelhante pelo mesmo sitio ou sistema de transporte. Sabe-se tambm que embora tais sistemas so
94
bastante eficientes para os L-aminocidos, o so muito pouco para as formas D, o que faz com que os D-aminocidos sejam pouco ou muito lentamente absorvidos. Os aminocidos que passam ao sangue portal e ao fgado so absorvidos no intestino delgado utilizando um mecanismo de transporte ativo que, na maioria dos casos, depende do sdio. A glicina, prolina e lisina no fogem do uso deste mecanismo para sua absoro, mas para eles no necessria a molcula de sdio. O transporte ativo, que um mecanismo de absoro mais rpido, requer de um transportador especfico. O transportador tem dois pontos de ligao especficos, um dos quais utilizado pelo nutriente a ser absorvido e o outro pelo sdio. O transportador carregado atravessa a membrana intestinal e deposita o nutriente e o sdio no interior da clula. Logo o transportador j sem o nutriente volta a atravessar a membrana para pegar mais outras molculas a serem transportadas. O sdio que ingressou clula bombeada ativamente ao lume do intestino, ficando disponvel para se ligar a outro transportador. Por sua vez os peptdeos e oligopeptdeos ingressam s clulas epiteliais do intestino delgado onde so hidrolisadas por di e tripeptidases especficas. 4. DIGESTO DOS COMPOSTOS NITROGENADOS NO INTESTINO GROSSO. Nos no-ruminates a maior parte dos produtos gerados pela digesto da protena no estmago e intestino delgado so absorvidos no intestino delgado. Ao ceco e colo dos no-ruminates normalmente chegam algumas formas de nitrognio das dietas que resistiram s aes qumicas enzimticas do estmago e intestino delgado e o nitrognio de origem endgena (secrees digestivas, clulas do trato digestivo, microorganismos). Nestes locais do intestino grosso no so produzidas enzimas e sim mucosas; consequentemente, a digesto sobre essas fontes de nitrognio devida s enzimas do intestino delgado (que passaram com o alimento ao ceco, e prolongam a sua atividade por quanto encontram um pH favorvel para tanto) e, principalmente, pelas fermentaes microbianas. Nos no-ruminantes a populao microbiana do intestino grosso , em conjunto, semelhante com a do rume dos bovinos; mas, a sua eficincia nos processos fermentativos sobre as fontes de nitrognio mais limitada seja porque o ceco e colo so menos desenvolvidos (aves, coelhos e sunos), seja porque pode ser rpida a passagem do alimento por estes locais, seja porque se apresentam em zonas que esto aps das regies de intensa absoro dos produtos gerados. Nos no-ruminantes os microorganismos degradam o nitrognio at aminocidos livres e NH3 que logo sero utilizados para a sntese de protena microbiana ou possivelmente absorvidos atravs das paredes do intestino grosso. A discusso que se apresenta na literatura est centralizada em estabelecer tanto a magnitude da absoro dos aminocidos quanto a sua participao no acrscimo de nitrognio s clulas do animal hospedeiro. O intestino grosso dos aves possui dois cecos, diferindo do dos mamferos que possuem um; contudo, este muito pequeno (5-10cm) e, em geral, tem um papel
95
bastante restrito quanto utilizao digestiva das fontes de nitrognio, sejam estas ligadas protena ou como nitrognio no protico. Nos coelhos o intestino grosso um local bastante volumoso, medindo cerca de 40 cm e com capacidade de aproximadamente 600 ml. A mucosa do ceco bem vascularizada e rica em clulas mucoprodutoras e de absoro. A poro proximal do ceco se relaciona com a juno leocecoclica, bastante importante na fisiologia deste e do clon. A poro distal do ceco apresenta um apndice, de 13 cm de comprimento em coelhos de 4 meses de idade, que contem numerosas clulas linfides (relacionadas com a secreo de ons bicarbonato, tamponantes dos cidos graxos volteis produzidos durante a fermentao cecal). neste apndice onde ocorre a fagocitose bacteriana e possivelmente a sntese de vitaminas hidrosolveis porquanto a apendicectomia reduz, significativamente, os nveis de vitamina B12 do contedo cecal. O clon tem um tamanho aproximado de 130 cm e est dividido em quatro regies: clon anterior proximal (5-15 cm), clon posterior proximal (de maior comprimento que o anterior), fusus coli (com comprimento muito pequeno, de 3 a 4 cm, este stio responsvel pela separao das fezes duras e moles) e clon distal (apresenta nmero elevado de clulas mucoprodutoras). Aps a digesto dos nutrientes do alimento no intestino delgado, os resduos ainda no digeridos passam pela vlvula leo-cecal distribuindo-se entre o ceco (a maior parte) e o clon proximal. O ceco tem importante papel na digesto do coelho devido fermentao que ali acontece, excreo seletiva de fibra e reingesto do contedo cecal (cecotrofia). No contedo cecal destacam-se as bactrias anaerbias, especialmente os bacilos no esporulados gram positivos, assim como a falta de lactobacilos, alm do mais neste local no existe uma populao significativa de protozorios provavelmente devido ausncia de substratos adequados como amido e acares solveis. No ceco as principais fontes de nitrognio para as bactrias so as que acompanham a digesta e a uria que vem do leo ou que difundida desde o sangue. Essas fontes de nitrognio so utilizadas pelas bactrias cecais na sntese de protena o que explicaria porque a porcentagem de aminocidos do contedo cecal superior da dieta. Todavia, s uma parte pequena dos aminocidos sintetizados pode ser absorvida por difuso atravs das paredes do ceco e clon. A maior parte destes encontram-se formando a protena microbiana e s teriam alguma utilidade para o coelho caso que fossem reingeridas com as fezes cecotrofas. O clon proximal realiza fortes movimentos anti-peristlticos que impulsionam os fludos e as partculas pequenas ao ceco onde os dois tipos de fezes (moles e duras) se misturam homogeneamente graas a sua constante movimentao e parte do material cecal passa ao clon proximal, onde igualmente objeto da ao dos movimentos antiperistlticos. Isto significa que o contedo cecal se encontra tanto no ceco como na parte proximal do clon. Consequentemente, o contedo cecal rico em material solvel e em partculas de tamanho pequeno, entanto que o material menos mido e formado por partculas grandes avana at o clon posterior onde sero formadas as fezes duras. O material contido no clon perde gua e rapidamente eliminado em resposta a uma estimulao nervosa.
96
No coelho as fezes so classificadas como moles (cecotrofos) e duras. Os cecotrofos so produzidos depois que o contedo cecal foi submetido durante algumas horas ao das bactrias. Sua produo inicia-se em resposta passagem completa do contedo cecal pela vlvula ileal. Por sua vez as fezes duras so constitudas pelas partculas maiores e sua eliminao sempre precede contraes simples e amplas do ceco e clon proximal, com rpida movimentao destas atravs do clon distal e reto; consequentemente, o coelho capaz de excretar rpida e seletivamente a fibra da dieta, retendo no ceco por tempo prolongado as fraes solveis e as partculas menores dos alimentos. A estratgia de produzir dois tipos de fezes capacita ao coelho a utilizar dietas altas em forragens, com parede celular de baixa digestibilidade e simultaneamente utilizar os demais constituintes das forragens. Quando a rea de maior atividade microbiana est localizada na parte posterior do trato gastrointestinal e logo dos mais importantes segmentos de atividade enzimtica e de absoro dos nutrientes, os microorganismos dispem de um substrato menos rico em nutrientes y energia disponveis. Por outro lado, embora h processos de inquestionvel valor nutricional para o hospedeiro como a absoro de AGV, a capacidade de utilizar outros produtos gerados pela atividade bacteriana (protena microbiana) limitada porque so limitados os sistemas de degradao enzimtica da protena e os de absoro dos produtos gerados. No coelho estas limitaes so compensadas com a cecotrofia que permite a digesto enzimtica das bactrias cecais, o aumento da digestibilidade da protena e o acrescimo da absoro intestinal dos aminocidos procedentes da protena bacteriana. Embora se sabe que no intestino grosso dos sunos ocorre intensa atividade de sntese e degradao microbiana dos aminocidos, a qual, alis, influencia a composio das fezes, h pouca evidncia que a mucosa do intestino grosso seja capaz de transportar os aminocidos e, portanto, so poucos os estudos que tm conseguido demonstrar que h absoro significativa deles neste local. Alguns pesquisadores encontraram, por exemplo, que a prolina, glicina, alanina, isoleucina, leucina, fenilalanina, metionina e triptofano no foram absorvidos no ceco isolado de sunos em crescimento e que os outros aminocidos s foram absorvidos em quantidades sem importncia. Experimentos realizados utilizando a tcnica da infuso de fontes proticas na regio distal do leo e no ceco, demostraram que a protena, os peptdeos e os aminocidos livres, bem de origem endgena ou do alimento, que atingem o intestino grosso, so degradados a aminocidos e amnia e incorporados s protenas microbianas, explicando deste modo uma alta parcela do nitrognio achado nas fezes. A amnia pode tambm ser absorvida e excretada como uria, quase de maneira completa e rpida na urina, sem se reter nos tecidos corporais, como pode ser observado nos resultados registrados na tabela 3. TABELA 3. EFEITO DA PROTENA FORNECIDA NA DIETA OU INFUNDIDA NO LEO TERMINAL SOBRE A EXCREO E A RETENO DE NITROGNIO (G/DA). Ingesto de Nitrognio Nitrognio fecal Nitrognio na urina Total RLP 1,00 1,42 3,31 CFR 17,34 1,83 5,89 CHEID 17,82 1,64 5,56 CHEIIT 17,55 1,50 17,32
97
Uria 1,64 3,78 3,00 15,88 Reteno de Nitrognio 0,73 0,05 (% do suplemento) Fonte: Fuller & Wang (1990) RLP: Rao livre de nitrognio CFR: Casena na rao CHEID: Casena hidrolizada com enzimas e infundida no duodeno CHEIIT: Casena hidrolizada com enzimas e infundida no leo terminal. No caso dos aminocidos por sua vez, os resultados da tabela 4, obtidos de vrias pesquisas, permitem estabelecer que a infuso de lisina no leo do suno no aumentou a reteno de nitrognio quando comparada com dietas basais deficientes neste aminocido. TABELA 4. EFEITO SOBRE A RETENO DE NITROGNIO DA SUPLEMENTAO OU DA INFUSO DE LISINA NO LEO TERMINAL DE SUNOS ALIMENTADOS COM UMA DIETA BASAL DEFICIENTE EM LISINA. Nvel de lisina Suplementao na dieta basal com lisina na dieta Lisina fornecida 4,9 13,6 (g/da) N retido (g/da) 6,4 14,6 Lisina infundida no elo terminal 13,8 6,6
Os resultados desses experimentos, conduziram a assinalar a inmeros pesquisadores que a protena e os aminocidos no intestino grosso so de pouco ou nenhum valor nutricional para os sunos porquanto no fazem aportes importantes de nitrognio para a sntese protica. Um outro fato marcante, ainda em discusso, est associado aos achados segundo os quais os aminocidos procedentes do alimento e os de origem endgena sofrem mudanas importantes na sua passagem pelo trato digestivo, sendo muito mais importantes os que se apresentam no intestino grosso. Finalmente, segundo pesquisadores dinamarqueses a protena retida e a resposta animal esteve mais associada com a digestibilidade da protena e dos aminocidos no leo terminal que com a digestibilidade fecal. Por sua parte, os pesquisadores franceses sugeriram que a digestibilidade ileal, quando comparada com a fecal, revela muito melhor as diferenas tanto entre as dietas como entre as fontes de protena, mas de maneira especial, quando elas so de baixa digestibilidade. Entretanto, o mesmos pesquisadores registraram que a digestibilidade ileal aparente dos aminocidos foi um critrio bem mais sensvel para revelar a existncia dos efeitos negativos do aquecimento do farelo de soja. 5. DESTINO DOS AMINOCIDOS ABSORVIDOS. O animal tem capacidade muito limitada de estocagem de aminocidos livres, somente o fazem por poucas horas; aps isso, os aminocidos podem tomar estes destinos: So utilizados para a sntese protica;
98
So utilizados para a sntese de aminocidos no essenciais, mediante transaminao; Servem como precursores de compostos nitrogenados: cidos nuclicos, creatina, colina e tiroxina; So utilizados para a sntese de glicose (gliconeognese). Participam deste processo todos os aminocidos no essenciais e por isso so chamados de aminocidos glicognicos; So convertidos a gordura (cetona e corpos cetnicos). Por esta razo so chamados de aminocidos cetognicos; Alguns aminocidos podem dar origem tanto a glicose (glicognicos) como cetona e corpos cetnicos (cetognicos); So deaminados, com o esqueleto carbonado sendo degradado a CO2, H2O e energia e o radical nitrogenado transformado em uria ou cido rico. O uso da protena e os aminocidos como fontes de energia produz aumento de calor entre 20 e 40%. No organismo animal parte da amnia empregada na sntese de aminocidos e outros compostos nitrogenados, mas sempre h um composto que precisa ser eliminado. A amnia liberada na degradao oxidativa dos aminocidos no pode estar em excesso no sangue do animal porquanto txica e at produzir a morte; para tanto deve ser convertida em compostos menos txicos. Na maioria dos mamferos, anfbios e peixes a amnia convertida no fgado e rins em uria e excretada pela urina. Nas aves e rpteis a amnia convertida em cido rico mediante um processo mais complexo. No primeiro grupo a uria sintetizada a partir de todos os radicais nitrogenados circulantes, mediante o denominado ciclo da uria. Clculos realizados por vrios pesquisadores permitem assinalar que o custo energtico da sntese da uria de 2 mol de ATP para cada tomo de nitrognio formado. Nas aves e repteis no existe o ciclo da uria, sendo que o processo de formao do cido rico mais complexo. Entanto muita pouca a excreo de L-aminocidos ntegros pela urina, j que a reabsoro renal muito eficiente, no ocorre o mesmo para os D-aminocidos. Para que os animais sintetizem suas protenas necessitam a presena simultnea de entre 18 e 22 aminocidos. Visto que alguns dos aminocidos no so sintetizados pelo organismo ou so a uma velocidade que no corresponde s necessidades do animal para o seu estado fisiolgico, idade e nvel de produo, devem ser fornecidos com a dieta. Esses so chamados aminocidos essenciais ou indispensveis. Aqueles outros aminocidos que no se encontram nestas condies, so conhecidos como aminocidos no essenciais ou dispensveis. Existe outro grupo de aminocidos conhecido como semi-indispensvel. Este grupo est formado por aminocidos no essenciais que podem sintetizar-se a partir de outro essencial, como o caso da cistina e tirosina formadas a partir da metionina e fenilalanina, respectivamente. Portanto, na formulao das dietas animais deve-se considerar a adio de metionina+cistina e fenilalanina+tirosina. A classificao dos aminocidos em essenciais, no essenciais e semi-indispensveis se baseia em consideraes de ordem metablica e depende, como j relatado, da espcie animal, do estado fisiolgico e do nvel de produo animal. No caso dos pintos e os sunos os aminocidos essenciais so: arginina, fenilalanina, histidina, isoleucina, leucina, lisina, metionina, treonina, triptofano, e valina. A glicina e a
99
serina no so considerados essenciais para as aves; entretanto, para as aves jovens, mais que nos adultos, a glicina deve ser adicionada dieta, pois este aminocido no sintetizado nos animais jovens a velocidade que estas necessitam para o crescimento rpido. Os nveis de serina e glicina so apresentados em conjunto em virtude do processo de interconverso entre estes dois aminocidos no ciclo da uria. Como a cistina tem como precursor a metionina, e a tirosina tem como precursor a fenilalanina, na formulao das dietas deve-se considerar a necessidade de metionina em separado e a de metionina-cistina em conjunto; situao similar acontece com a fenilalanina-tirosina. Em alimentao animal, se um aminocido tem um nico precursor essencial, se considera tambm como aminocido essencial a soma dos dois. Na literatura sobre nutrio animal tambm relatado o termo aminocido limitante para descrever aquele ou aqueles aminocidos essenciais que por no estar presente nas dietas em quantidades suficientes limita o crescimento ou a produo animal, ainda quando haja excesso de todos os outros aminocidos. Espera-se que sua adio extra s dietas elimina estes problemas. Nas dietas prticas para aves e sunos a lisina e a metionina so tidos geralmente como os primeiros aminocidos limitantes, visto que esto em baixa concentrao na maior parte das protenas dos alimentos freqentemente utilizados e tambm porque elas so susceptveis formao da reao de Maillard. Quando h desproporo entre L-aminocidos de estrutura semelhante, aquele de menor teor pode ter sua excreo urinria aumentada. Este fenmeno conhecido na literatura como antagonismo entre aminocidos. Um exemplo clssico de antagonismo verificado na alimentao animal o encontrado entre a lisina e a arginina. Quando h excesso de lisina na dieta, ocorre alta excreo de arginina, aps seu desdobramento a ornitina e uria. O antagonismo entre ambos os aminocidos explicado porque a lisina compete com a arginina pela reabsoro nos tbulos renais, alm de que o excesso do primeiro aumenta a atividade da arginase renal com o que se incrementa o catabolismo e destruio do sedundo. Outros casos de antagonismo entre aminocidos podem se apresenta com o excesso na dieta de isoleucina, histidina e tirosina e vice-versa. Alm do antagonismo tambm so registrados ocorrncias de toxicidade quando o efeito adverso do excesso de um aminocido no pode ser superado ao fornecer o seu aminocido antagnico. Um exemplo clssico deste tipo de situao se apresenta com o excesso de metionina sinttica decorrente de uma dosagem incorreta do aminocido. Na formulao de uma dieta necessrio levar em considerao os conceitos de aminocido essencial, no essencial, semi-indispensvel, limitante, antagonismo, toxicidade e desequilbrio de aminocidos. Tanto o excesso quanto a falta de aminocidos nas dietas so prejudiciais por romperem o equilbrio dinmico do metabolismo. O desequilbrio se d quando adieta no est balanceada em relao aos aminocidos presentes ou quando o teor de protena inferior ao necessrio. A deficincia de um s aminocido essencial, impede a realizao da sntese protica; portanto, a insuficincia de aminocidos individuais favorece a deaminao dos demais aminocidos, a perda de amna como uria e o consumo da cadeia de carbono na obteno de energia.
100
O problema de desequilbrio de aminocidos resolvido pela adio de quantidades extras de aminocidos limitantes ou desequilibrando o teor desses aminocidos mediante a combinao de alimentos. Um exemplo clssico de combinao entre alimentos encontra-se entre o milho e o farelo de soja. Entanto o primeiro tem baixos teores de lisina e triptofano, o que causa o crescimento muito lento do animal, o farelo de soja apresenta quantidades adequadas de lisina e triptofano, porm deficiente em metionina e cistina e sozinha, portanto, no pode apoiar o crescimento normal dos animais. Quando o milho se combina com o farelo de soja se complementem mutuamente ao compensarem suas deficincias individuais de aminocidos. REFERNCIAS BIBLIOGRFICAS. 1. Batterham. E.S. Availability and utilization of amino acids for growing pigs. Nutrition Research Reviews. 5: 1-18. 1992. 2. Christensen, H.N. Amino acid nutrition: A two-step absortive process. Nutrition Reviews. 51 (4): 95-100. 1993. 3. Christensen, H.N. What is the physiological origin of free D-amino acid in mammals. Nutrition Review. 50 (10): 294-295. 1992. 4. Chung, T.K.; Baker, D.H. Utilization of methionine isomers and analogs by the pig. Canadian Journal of Animal Science. 72 (1): 185-188. 1992. 5. Collarini, E.J.; Oxender, D.L. Mechanisms of the transport of amino acids across memebranes. Annual Review Nutrition. 7: 75-90. 1987. 6. Darragh, A.J. et al. Absorption of lysine and methionine from the proximal colon of the piglet. British Journal of Nutrition. 71 (5): 739-752. 1994. 7. Gardner, M.L.G. Gastrointestinal absorption of intact proteins. Annual Review Nutrition. 8: 329-350. 1987. 8. Harper, A.E. Nonessentialamino acids. Journal of Nutrition. 104: 965-967. 1974. 9. Hintz, H.F.; Cymbaluk, N.F. Nutrition of horse. Annual Review of Nutrition. 14: 243-268. 1994. 10. Just, A. The digestive capacity of the caecum-colon and the value of the nitrogen absorbed from the hind gut for protein synthesis in pigs. British Journal of Nutrition. 46 (1): 209-219. 1981. 11. Kye-Wing, C.; MacKenzie, W. Substitution of five essential amino acids by their alfa-ketoanalogues in the diet for rats. Journal of Nutrition. 104: 1208-1214. 1974. 12. Man, E.H. Dietary D-amino acids. Annual Review Nutrition. 7: 209-225. 1987. 13. Rrat, A. et al. Absorption kinetics of dietary hydrolysis products in conscious pigs given diets with different amount of fish protein. Amino-N and glucose. British Journal of Nutrition. 60: 91-104. 1988. Individual amino acids. British Journal of Nutrition. 60: 105-120. 1988. 14. Tesseraud, S. Mtabolisme protique chez le poulet en croissance. Effet des protines alimentaires. Productions Animales. 8 (3): 197-212. 1995. 15. Tyler, H.D. et al. Developmental appearance of peptide-binding sites in the small intestine of the neonatal piglet. Canadian Journal of Animal Science. 74 (2): 243-250. 1994. 16. Webb, K.E. et al. Peptide absorption: A review of current concepts and future perspectives. Journal of Animal Science. 70 (10): 3248-3257. 1992. 17. Webb, K.E. Intestinal absorption of protein hydrolysis products: A review. Journal Animal Science. 68 (9): 3011-3022. 1990.
101
MTODOS DE AVALIAO DA DISPONIBILIDADE DA PROTENA E DOS AMINOCIDOS NOS ALIMENTOS PARA NO RUMINANTES. 1. INTRODUO. No decurso dos dois ltimos sculos, os clculos de necessidades de alimentos para os animais domsticos e o planejamento das dietas tm sido baseados em padres de alimentao, nos quais so expressos os requisitos dos animais bem como o valor nutritivo dos alimentos (Flatt, 1988). Para a nutrio isto tem significado que no importante estimar os requisitos dos animais em termos das necessidades metablicas se no existe informao do valor potencial dos alimentos expressa nas mesmas unidades (Fuller, 1988); mas tambm, esta metodologia tem demonstrado que muitos dos erros associados avaliao dos alimentos esto compensados, de certo modo, pelas maneiras como so realizadas as estimativas dos requisitos animais (Flatt, 1988). Para a nutrio animal, todos os nutrientes so importantes, mesmo que sejam requeridos em pequenas quantidades, mas a avaliao dos alimentos tem sido orientada energia, que representa o alimento como um todo, e ao contedo de protena, mas principalmente os aminocidos essenciais, como parte da matria orgnica, porque os dois so os componentes mais importantes das dietas em termos quantitativos (Bickel, 1988). No que diz a respeito dos sunos, por muitos anos os padres de alimentao da Agricultural Research Council (ARC, 1978) e da National Research Council (NRC, 1988) tm utilizado os clculos dos requisitos nutricionais e as estimativas do valor dos alimentos baseados nos contedos totais de protena e de alguns aminocidos essenciais No entanto, Mitchell (1924) j havia alertado para a necessidade de se referir protena lquida de um alimento como uma forma de expressar seu verdadeiro valor, expresso que mudou mais tarde quando o mesmo Mitchell, em 1964, citado por Sibbald (1987), observou que a avaliao nutricional de um alimento estava associada s diferenas na disponibilidade de seus aminocidos. Foi, ento, baseados no conceito de disponibilidade, mais que no contedo total de protena e aminocidos, que comearam a surgir, a partir dos anos oitenta, as novas propostas de avaliao dos alimentos e de estimativas dos requisitos nutricionais no s para os sunos mas tambm para as aves. Como manifestou Sibbald (1987), com a adoo deste critrio, espera-se melhorar a formulao das raes, dar mais ateno aos efeitos dos excessos e desbalanceamentos, mudar o objetivo dos padres de alimentao atuais que visam mais suprir as deficincias e, alis, poderiam se diminuir os nveis de incorporao da protena s dietas, provocando, possivelmente, reduo nos custos de alimentao. Segundo Durmad et al. (1995), a adoo do critrio de disponibilidade dos nutrientes no clculo das dietas trar uma diminuio na excreo de nitrognio com a conseqente reduo na poluio da gua por nitritos e do ar por emisses de amnia. Estes aspectos esto estimulando a realizao de estudos que visam desenvolver novas metodologias de avaliao nutricional e estimativas das exigncias dos animais
102
baseadas na disponibilidade da protena e dos aminocidos. No obstante, para Bellaver (1989), estas novas propostas se enfrentam a fortes presses, as quais esto precisamente no fato de que elas s sero teis produo de alimentos se as frmulas baseadas em nutrientes disponveis promoverem diferenas no desempenho animal quando comparadas quelas elaboradas a partir de contedos totais. Como foi citado anteriormente, qualquer padro de alimentao possui dois componentes nutricionais bsicos que devem ser estabelecidos simultaneamente e expressos nas mesmas unidades de medida: a informao do valor nutricional dos alimentos e as estimativas das exigncias dos animais. O objetivo central deste trabalho foi estudar os fundamentos de alguns dos principais mtodos e tcnicas que existem para avaliar a disponibilidade da protena e os aminocidos dos alimentos para sunos, por quanto este componente oferece grandes possibilidades de se pesquisar com sucesso nas nossas condies. Importante salientar que apesar da maior parte das pesquisas estudadas foram realizadas em sunos, as metodologias utilizadas nos diferentes experimentos e muitos de seus achados podem servir como referncia para os outros no-ruminantes. 2. CARACTERSTICAS GERAIS DAS PROPOSTAS DE AVALIAO DA DISPONIBILIDADE DA PROTENA E OS AMINOCIDOS DOS ALIMENTOS PARA NO RUMINANTES.
O primeiro fato marcante que existe na avaliao da protena dos alimentos para no ruminantes que muitos dos mtodos clssicos descritos na literatura foram realizados com pequenos animais, principalmente ratos e aves, pensando-se nas necessidades de desenvolver estratgias de avaliao que fossem de utilidade nos humanos (Flatt, 1988). Destes mtodos fazem parte, de acordo com Fuller (1988), a relao de eficincia protica, o valor biolgico da protena, o ndice de balano de nitrognio, e a utilizao lquida da protena. Ainda que muitos destes no sejam frequentemente empregados nos alimentos fornecidos aos sunos, tm provisto alguns de seus princpios mais importantes para se utilizar nos estudos de disponibilidade da lisina, metionina, treonina e triptofano realizados na Austrlia por Batterham et al.(1990), Beech et al.(1991) e Batterham (1992). O segundo tema a destacar que existem vrios experimentos isolados onde tm sido testados inmeros mtodos e tcnicas de avaliao nutricional, porm no fcil achar na bibliografia uma proposta unificada de avaliao da disponibilidade da protena e aminocidos; ao respeito, Bellaver (1989) indagou sobre a necessidade de padronizar as diferentes propostas e procedimentos de avaliao com vistas a ter comparaes vlidas e enriquecer as tabelas de composio de alimentos, bem como incluir nestas a determinao das exigncias dos nutrientes disponveis. Da variada literatura consultada s foi possvel identificar quatro propostas que apresentaram um corpo coerente de idias sobre o significado do valor nutricional das protenas e dos aminocidos para sunos. A tabela 1 registra um resumo das caractersticas mais marcantes que apresentam cada uma delas.
103
TABELA 1. PROPOSTAS DE AVALIAO DA DISPONIBILIDADE DA PROTENA E OS AMINOCIDOS DOS ALIMENTOS. AUTOR MTODOS in vivo TCNICAS Digestibilidade ileal dos AA MTODOS in vitro TCNICAS Disponibilidade dos grupos livres da lisina (FDNB) Liberao de AA com incubaes enzimticas
Excreo fecal dos AA ou o Nitrognio (N) ZEBROWSKA (1978) Concentrao de AA no sangue Ensaios de crescimento Avaliaes da disponibilidade dos AA DIRETOS Ensaios de crescimento Experimentos de balano Ensaios com sacos de nilon SIBBALD (1987) INDIRETOS Ensaios com insetos Ensaios microbiolgicos Determinaes de AA no plasma NVEL DIGESTIVO Desaparecimento no trato gastrointestinal Aparecimento dos nutrientes no sangue HENRY (1985) NVEL METABLICO Reteno de N e AA Utilizao Lquida da Protena (NPU) Ensaios de crescimento Sacrifcios comparativos INTEGRAIS Relao de Eficincia Protica (PER) ndice Lquido da Protena (LPI) ndice de Crescimento
Digestes enzimticas Prova com hidroxiprolina Composio qumica do alimento
104
do Nitrognio (ICN) Utilizao Lquida da Protena (NPU) Valor Biolgico (VB) FULLER (1988) ADITIVOS Concentro de AA no alimento Digestibilidade (Absoro) Ensaios de crescimento (Disponibilidade) A primeira proposta a ser apresentada a formulada por Zebrowska (1978). Esta pesquisadora destacou que os mtodos para estimar a disponibilidade podem classificar- se segundo sejam in vivo e in vitro, cada um dos quais possuindo diferentes tcnicas. Do primeiro grupo fazem parte os ensaios de crescimento, as dosagens da concentrao de aminocidos livres no sangue, a excreo fecal destes ou de nitrognio e as avaliaes de disponibilidade e digestibilidade ileal. Nos mtodos in vitro, encontram-se as determinaes colorimtricas dos grupos livres da lisina e as taxas de liberao de alguns aminocidos das protenas submetidas a diferentes incubaes enzimticas. Sibbald (1987) compartilhou com Zebrowska (1978) a idia de que a disponibilidade da protena e dos aminocidos para sunos e aves pode ser bem estimada com mtodos in vivo ou in vitro. Mas este pesquisador canadense acrescentou o nmero de tcnicas a serem levadas em considerao para cada grupo de mtodos, alm de incluir os ensaios de degradabilidade com sacos de nilon mveis no mesmo grupo de experimentos de balano e de crescimento, apesar de que para diversos autores eles fazem parte de uma das modalidades dos estudos de digestibilidade (Sauer et al., 1989; Leibholz, 1991; Koelen et al., 1992) ou so uma proposta intermediria entre os mtodos in vivo e in vitro (Boisen & Eggum, 1991). Na proposta Sibbald (1987) estabeleceu que as determinaes in vivo devem dividir-se em mtodos diretos e indiretos. No primeiro grupo esto os ensaios de crescimento, os experimentos de balano e os ensaios com sacos de nilon. No entanto no grupo dos mtodos indiretos, encontram-se os ensaios com insetos, os microbiolgicos, as determinaes de aminocidos no plasma e as tcnicas de digestibilidade. Finalmente, as determinaes in vitro so realizadas utilizando as digestes enzimticas, o mtodo da hidroxiprolina ou levando em considerao a composio qumica como mtodo para estimar o valor da protena. Henry (1985) e Fuller (1988), por sua vez, formularam propostas que se distanciam das anteriores pelo fato de que nelas no so includos os mtodos in vitro como parte das estratgias de avaliao, mas, quando so comparadas, mostram que entre elas h diferenas marcantes a serem destacadas. Assim, por exemplo, Fuller (1988) estabeleceu que os mtodos de avaliao podiam ser chamados de integrais e aditivos. Nos primeiros, a protena tem um valor nico e no aditivo, o qual representa sua capacidade global de afetar a taxa de crescimento ou a reteno de nitrognio, sem levar em considerao as necessidades deste elemento para o metabolismo basal e sem fornecer informao sobre os fatores que determinaram este valor. Destes mtodos fazem parte tcnicas tais como a relao de eficincia protica (PER), relao lquida de protena (NPR), ndice de crescimento de nitrognio, utilizao liquida de
105
protena (NPU) e o valor biolgico (VB). Para Fuller (1988), nos mtodos aditivos a avaliao de uma protena surge da estimativa da quantidade de cada um dos aminocidos de importncia nutricional que so fornecidos pelo alimento de uma forma tal que sejam absorvidos e utilizados pelo animal. Nestes, existem trs componentes que devem ser conhecidos para avaliar uma protena: a concentrao do aminocido no alimento, a frao que absorvida, ou seja, sua digestibilidade, e desta, a frao que e utilizvel, isto , sua disponibilidade, a qual avaliada nos ensaios de crescimento. Entretanto, Henry (1985) promoveu a idia de que a avaliao da protena atingiria dois nveis: o desaparecimento no trato gastrointestinal e, portanto, seu aparecimento no sangue, e o nvel metablico com as tcnicas prprias dos estudos de reteno de nitrognio e de aminocidos limitantes, como so a NPU, os ensaios de crescimento e o sacrifcio comparativo. O ltimo assunto a destacar nas diferentes propostas de avaliao a existncia de vrias maneiras de se interpretar o termo disponibilidade. No entanto este termo s vezes utilizado de maneira semelhante, contudo apresenta considerveis diferenas quanto seu significado. 3. MTODOS PARA EXPRESSAR A DISPONIBILIDADE DA PROTENA E OS AMINOCIDOS DOS ALIMENTOS. 3.1. OS ESTUDOS DE AVALIAO in vivo E O CONCEITO DE DISPONIBILIDADE. No contexto do animal a disponibilidade interpretada de maneira diferente daquela como entendida nas metodologias in vitro. Nos animais a digestibilidade e a disponibilidade definem o valor potencial da protena dos alimentos (Fuller & Wang, 1990) e mesmo que os dois termos estejam relacionados e tenham significados diferentes, frequente achar estudos onde so utilizados como sinnimos, o que, de acordo com Batterham (1992), este fato explicado pela crena de que se um nutriente digerido, est em forma disponvel para ser usado. Sibbald (1987) esclareceu perfeitamente a relao entre os dois termos ao assinalar que a absoro dos nutrientes no trato digestivo pr-requisito para a sua utilizao, mas no demonstra que tenha que existir disponibilidade: alguns aminocidos presentes por exemplo nas protenas danificadas pelo calor podem ser absorvidos e excretados na urina, o que, em conseqncia, indicaria que no se encontraram disponveis para serem utilizados no metabolismo animal. O memo Sibbald (1987) considerou, por um outro lado que a utilizao de um nutriente que foi absorvido no trato evidncia da sua disponibilidade, porm, a excreo no prova a perda da mesma. A digestibilidade, ento, deve ser entendida como a definiu Rrat (1990): uma medida do desaparecimento da protena e dos aminocidos durante sua passagem pelo sistema digestivo e no sob a idia de absoro, como tem sido adotada por muitos investigadores, visto que estas fontes de nitrognio podem ser destrudas e modificadas pela ao dos microrganismos em alguns locais do trato ou metabolizadas pelas paredes do mesmo durante a absoro. Por sua vez, a absoro refere-se aos intercmbios que acontecem entre o sangue e o lume digestivo, os quais so medidos de maneira simultnea pelas diferenas entre as concentraes dos nutrientes no sangue portal e arterial e o fluxo de sangue na veia porta (Rrat, 1990). No entanto, a disponibilidade metablica deve ser definida segundo como a entendeu Batterham (1992): a proporo daquele aminocido limitante da dieta que foi digerido e absorvido e que utilizado para a sntese de protena.
106
As diferenas conceituais entre estes termos levam, ento, a considerar a necessidade de se analizar algumas das metodologias e tcnicas que h para realizar as medies certas do desaparecimento, absoro e disponibilidade da protena e os aminocidos, mas tambm obriga conhecer as caractersticas dos tipos de associao entre a digestibilidade e a disponibilidade metablica destes nutrientes. 3.1.1. A digestibilidade como estratgia de avaliao da protena e dos aminocidos. 3.1.1.1. Digestibilidade fecal aparente e fecal verdadeira. Tal vez as bases metodolgicas dos experimentos sobre digestibilidade que foram realizados em sunos e ratos por Dammers (1964) e Eggum (1973), citados por Lenis (1992), estejam no trabalho clssico desenvolvido em ratos por Kuiken & Lyman (1948). Nelle foi sugerido o uso do mtodo de medio do ndice fecal corrigido pelo contedo dos componentes nitrogenados de origem endgena (secrees digestivas, liberao de clulas do trato gastrointestinal e microrganismos) para determinar a digestibilidade fecal verdadeira. Este tipo de medio diferente da aparente porquanto nesta ltima no so tidas em considerao as correes dos aportes endgenos de compostos nitrogenados incorporados s fezes (Zebrowska ,1978; Low, 1982; Henry, 1985; Sauer & Ozimek, 1986). De acordo com relatos de Henry (1985), so mais altos os valores estimados da digestibilidade fecal verdadeira dos aminocidos que os obtidos com a metodologia aparente. Mas, para as condies prticas de avaliao da protena, o mesmo pesquisador reconheceu que no existe vantagem em usar a digestibilidade verdadeira, pois os padres de aminocidos digestveis so muito similares nas duas tcnicas, sendo que os mtodos existentes para a correo do nitrognio endgeno tm limitaes reais e, ademais, as perdas endgenas, como parte dos requisitos para mantena, so consideradas na digestibilidade aparente. Neste ponto, importante destacar as sugestes de Darcy-Vrillon & Rrat (1983), citados por Henry (1985), segundo as quais, com as dietas completas, o mais importante conhecer as quantidades totais de aminocidos fornecidos pelo intestino, independente de serem de origem endgena ou do alimento. No caso dos alimentos individuais, a estratgia sugerida que seja considerada a possibilidade de estimar a frao endgena e, consequentemente, impretervel a determinao da digestibilidade verdadeira. 3.1.1.2. Digestibilidade ileal aparente e ileal verdadeira. Se bem durante quase duas dcadas foi aceitada a digestibilidade fecal como proposta metodolgica, os estudos desenvolvidos a partir dos anos setenta por vrios grupos de pesquisadores e sob as mais diversas condies, mostraram que a digestibilidade ileal dos aminocidos melhor estimativa do valor nutricional das protenas que sua digestibilidade ileal e, em consequncia, um mtodo mais til para se incluir na formulao das dietas para sunos (Just et al., 1985; Darcy-Vrillon & Laplace, 1990; Fuller & Wang, 1990; Moughan et al., 1991; Bellaver, 1994; Williams, 1995) e aves (Raharjo & Farrel, 1984/1985; Papadopoulos, 1985; Parsons, 1990; Johnson, 1992; Rostagno et al., 1995). Esta virada nos estudos de digestibilidade pode ser explicada pelos achados das pesquisas na rea da fisiologia digestiva e o metabolismo dos compostos nitrogenados, alguns dos quais sero relatados a seguir:
107
Se bem que se saiba que no intestino grosso dos sunos ocorre intensa atividade de sntese e degradao microbiana dos aminocidos, a qual, alis, influencia a composio das fezes (Mason, 1984), h pouca evidncia que a mucosa do intestino grosso seja capaz de transportar os aminocidos e, portanto, so poucos os estudos que tm conseguido demonstrar que h absoro significativa deles neste local (Zebrowska, 1978). Por exemplo, Olszewski (1975), citado por Sauer & Ozimek, 1986), encontrou que a prolina, glicina, alanina, isoleucina, leucina, fenilalanina, metionina e triptofano no foram absorbidos no ceco isolado de sunos em crescimento, no entanto que os outros aminocidos s foram absorbidos em quantidades sem importncia; Os experimentos pioneros de Zebrowska (1973), citados por Zebrowska (1978), utilizando a tcnica da infuso de fontes proticas na regio distal do leo e no ceco, demostraram que a protena, os peptdeos e os aminocidos livres, bem de origem endgena ou do alimento, que atingem o intestino grosso, so degradados a aminocidos e amnia e incorporados s protenas microbianas, explicando deste modo uma alta parcela do nitrognio achado nas fezes. A amnia pode tambm ser absorvida e excretada como uria, quase de maneira completa e rpida na urina, sem se reter nos tecidos corporais, como pode ser observado nos resultados registrados na tabela 2. TABELA 2. EFEITO DA PROTENA FORNECIDA NA DIETA OU INFUNDIDA NO LEO TERMINAL SOBRE A EXCREO DE NITROGNIO FECAL E NA URINA (g/dia) Ingesto de Nitrognio Nitrognio fecal Nitrognio na urina Total 3,31 Uria 1,64 Reteno de Nitrognio (% do suplemento) Fonte: Fuller & Wang (1990) RLP: Dieta livre de nitrognio CFR: Casena na dieta CHEID: Casena hidrolizada com enzimas e infundida no duodeno CHEIIT: Casena hidrolizada com enzimas e infundida no leo terminal Os resultados destes experimentos, corroborados mais tarde por Just et al. (1981), conduziram a assinalar que no intestino grosso a protena e os aminocidos so de pouco ou nenhum valor nutricional para os sunos porque no fazem aportes importantes de nitrognio para a sntese protica. Um outro fato marcante, ainda em discusso, est associado aos achados de Sauer & Ozimek (1986) e Lenis (1992), segundo os quais os aminocidos procedentes do alimento e os de origem endgena sofrem mudanas importantes em sua passagem RLP 1,00 1,42 CFR 17,34 1,83 5,89 3,78 0,73 CHEID 17,82 1,64 5,56 3,00 CHEIIT 17,55 1,50 17,32 15,88 0,05
108
pelo trato digestivo, sendo muito mais importantes os que se apresentam no intestino grosso; Finalmente, segundo Just et al. (1985), a protena retida e a resposta animal esteve mais associada com a digestibilidade da protena e dos aminocidos no leo terminal que com a digestibilidade fecal. Por sua parte Henry (1985) sugereu que a digestibilidade ileal, quando comparada com a fecal, revela muito melhor as diferenas tanto entre as dietas como entre as fontes de protena, mas de maneira especial, quando elas so de baixa digestibilidade. Entretanto, o mesmo pesquisador registrou que a digestibilidade ileal aparente dos aminocidos foi um critrio bem mais sensvel para revelar a existncia dos efeitos negativos do processamento calrico no farelo de soja. Diante desta mudana nas estratgias de avaliao nutricional dos alimentos, o estudo das metodologias de colheita da digesta ileal converteu-se, rapidamente, em fator chave das pesquisas que tm por objetivo estimar a digestibilidade ileal dos componentes nitrogenados em sunos. 3.1.1.2.1. Mtodos de colheita do contedo ileal para estimar a digestibilidade da protena e dos aminocidos. H vrios mtodos de colheita da digesta intestinal que j foram testados visando estimar a digestibilidade da protena e dos aminocidos em sunos; contudo, segundo Leeuwen et al. (1991), cada um deles deve cumprir com duas exigncias bsicas: No podem afetar as condies fisiolgicas gerais normais dos animais e As amostras colhidas devem representar as condies dos processos digestivos. Recentemente, Giraldo (1996) realizou uma avaliao crtica dos inmeros mtodos de colheita do contedo ileal quanto as vantagens, problemas e limitaes que oferecem, tanto com relao qualidade da digesta colhida como sobre a fisiologia digestiva e as condies gerais dos animais. A siguente uma proposta de classificao dos diferentes mtodos relatados na literatura para atingir este objetivo e usados visando estimar a digestibilidade ileal dos aminocidos: Mtodo do sacrifcio de animais. Sugerido para ser realizado em ratos (Kuiken & Lyman, 1948), ratos e sunos (Thorpe & Thomlinson, 1967; Moughan et al., 1984; Skilton et al., 1991; Donkoh et al., 1994 a,b,c) e em aves (Raharjo & Farrel, 1984/1985); Mtodos de colheita do quimo ileal com animais modificados cirurgicamente: Uso de cnulas simples em forma de T, colocadas no leo distal entre 5 e 15 cm antes do esfincter ileocecal em sunos de diferentes idades (Decuypere et al., 1977; Gargallo & Zimermann, 1980; Walker et al., 1986; Leewen et al., 1987, 1991), aves (Raharjo & Farrel, 1984/1985) e ainda em coelhos (Gidenne et al., 1994); Tcnica da cnula em forma de T, inserida no ceco aps do esfincter ileocecal, desenvolvida pela equipe comandada por Leeuwen et al. (1988) na Holanda; Uso de cnulas reentrantes: Tipo ileoileal (Cunningham et al., 1963) Ileocecal (Easter & Tanksley, 1973)
109
A cnula ps-valvular leo-clica, proposta por pesquisadores franceses (Darcy et al., 1980 a,b,c; Darcy-Vrillon & Laplace, 1985 a,b; 1990); A anastomose ileoretal, desenvolvida por vrios grupos de pesquisadores com a preservao ou no do esfncter ileocecal (Fuller et al., 1994; Laplace et al., 1994). Da mesma maneira que a fecal, a digestibilidade ileal pode ser expressa como verdadeira ou aparente, dependendo se so ou no consideradas as correes feitas pelos aportes endgenos dos compostos nitrogenados (Henry, 1985; Sauer & Ozimek, 1986; Fuller & Wang, 1990; Lenis, 1992; Williams, 1995). No intestino delgado, as secrees digestivas originrias da saliva, suco gstrico, bile e secrees pancreticas e intestinais contribuem com a maior parcela dos aportes endgenos de nitrognio digesta. Henry (1985) estimou que em sunos em crescimento estes aportes eram, sem considerar o suco gstrico, da ordem de 20 a 25 g de nitrognio por dia, representados, em sua maior parte, por aminocidos, uria e amnia. 3.1.1.3. Mtodos para estimar a frao endgena do nitrognio nos estudos de digestibilidade ileal e fecal verdadeira. Nos estudos de digestibilidade ileal ou fecal verdadeira existem pelo menos cinco mtodos para se estimar a frao endgena de nitrognio: Mtodo direto, como foi denominado por Sauer & Ozimek (1986), baseado na alimentao de grupos de animais com dietas livres de protena. Bem que seja a forma mais simples e fcil para estimar o nitrognio endgeno, este mtodo tem sido muito questionado porque o processo digestivo apresenta um comportamento diferente daquele que teria com o fornecimento de uma dieta normal; assim, por exemplo, Donkoh et al. (1995) relataram que as excrees encontram-se diminuidas com tais raes, o que segundo Rrat (1990) e Lenis (1992) resultaria numa subavaliao das perdas dos aminocidos endgenos totais. Alm disso, de acordo com Low (1982), no caso das dietas que so empregadas de maneira frequente na alimentao animal, o teor e o tipo de fibra aumentaro as secrees e o crescimento microbiano. Finalmente importante assinalar que em sunos ainda no h informao suficiente para se estimar os efeitos da durao deste tipo de alimentao sobre os valores de digestibilidade e as condies gerais dos animais, como sim acontece nos estudos de determinao da energia metabolizvel verdadeira (EMV) e a biodisponibilidade dos aminocidos utilizando a metodologia proposta por Sibbald (1987) para avaliar os alimentos oferecidos as aves. Uso de dietas semi-sintticas com diferentes nveis de casena lctica ou a fonte de protena a se testar. Neste caso, a frao endgena estimada quando o consumo de nitrognio zero, empregando uma anlise de regresso (Furuja & Kaji, 1989). Este mtodo foi questionado por Donkoh et al (1995) porque a estimativa das perdas endgenas de nitrognio so restritas a uma funo linear e visto que a tcnica da regresso assume que estas permanecem constantes independente dos acrescimos no teor de protena ingerida, a proposta nem sempre faz uma descrio real do fenmeno biolgico; Tcnicas de diluio de istopos. Desenvolvidas h muito tempo em pesquisas com ratos, nos ltimos cinco anos vm sendo includas nos experimentos com sunos (Moughan et al., 1992; Armstrong et al., 1995); no obstante, sejam propostas de custos muito mais altos, elevada complexidade nas medies (Low, 1982) e, ainda, s determinam o nitrognio total recuperado e no cada um dos aminocidos (De
110
Lange et al., 1989). Quando se introduz a correo do nitrognio endgeno utilizando estas tcnicas, a digestibilidade ileal ou fecal denominada de real (Jondreville, 1994), o que poderia explicar porque os pesquisadores consideram que ela oferece uma definio bem melhor da natureza dos compostos nitrogenados endgenos que aparecem no contedo ileal ou nas fezes; Donkoh et al. (1995) proposeram um novo mtodo para determinar a excreo ileal de aminocidos endgenos que consiste na alimentao do animal com peptdeos pequenos (com peso molecular menor a 5000) originados da hidrlise enzimtica da casena, seguida da ultrafiltrao e da medio do resduo. A caracterizao das fraes de alto peso molecular da digesta ileal que surgem aps da precipitao e a ultrafiltrao, pode ser considerada como uma medida das perdas dos aminocidos endgenos. O emprego deste tipo de alimentao mantem aparentemente normal os nveis de excreo do nitrognio endgeno. Segundo as pesquisas iniciais, considera-se que esta tcnica seja melhor que as propostas que utilizam as dietas livres de nitrognio e as equaes de regresso, porm os primeiros resultados sugerem que na digesta dos ratos o valor mdio do fluxo de aminocidos endgenos geralmente mais baixo, alis que s seja aplicvel na correo da determinao da digestibilidade das protenas de origem animal, as quais no tm fibra e fatores antinutricionais; Finalmente, existem tcnicas para estimar o nitrognio endgeno de origem microbiana baseadas no uso dos cidos desoxirribonuclico e diaminopimlico, considerados como marcadores da atividade microbiana e de freqente utilizao nos estudos de fisiologia digestiva nos ruminantes (Rowan et al., 1992). Por enquanto, estas propostas no possuem uma avaliao crtica de suas possibilidades e problemas quando utilizadas nas pesquisas com sunos. Uma questo adicional que deve ser abordada nos estudos de digestibilidade refere-se necessidade de que as amostras de digesta como de fezes colhidas refletem as condies de estabilidade dos processos digestivos. O problema que, pelo menos no caso da digestibilidade ileal, o fluxo digestivo neste local do trato gastrointestinal no contnuo, e, portanto, no possvel fazer uma colheita quantitativa do quimo (Low, 1980; Oslage et al., 1989). A soluo desta situao pode-se atengir introduzindo controles as quantidades, freqncia, apresentao e os horrios de fornecimento dos alimentos; mas tambm, o uso de indicadores inertes de utilidade como elemento que possibilita estimar se tal condio foi conseguida (Saha & Gilbreath, 1993). No que diz, respeito por exemplo dos experimentos de digestibilidade que empregam as cnulas simples em forma de T, inseridas no leo distal e no ceco aps do esfncter ileocecal, e os que se baseam na colheita total de fezes os pesquisadores sugerem que imprescindvel o uso dos indicadores; no entanto assinalam tambm que possvel acrescentar os problemas de preciso dos resultados, explicados pela distribuio e localizao no trato, as variaes na recuperao e o fato de que os indicadores nem sempre refletem as condies do processo digestivo (Klher et al., 1990). Para a soluo destes problemas Oslage et al.(1989) proposseram perodos de amostragem muito mais longos, no entanto que Leeuwen et al.(1987) sugereram o uso de uma mistura que tenha indicadores de fase slida e lquida. O problema que, como foi considerado por Thomas (1988), ainda existem dvidas em relao obteno de amostras representativas do comportamento que tm estes indicadores quanto sua passagem pelo trato gastrointestinal.
111
3.1.1.4. Digestibilidade ileal ou fecal: possvel uma escolha? Os estudos relatados por Henry (1985) mostraram que, no caso do nitrognio, geralmente os valores de digestibilidade fecal so, em mdia, mais altos que os da ileal, com variaes nas diferenas entre 2 e 14 pontos. Para este nutriente o mesmo autor considerou que ainda continua sendo importante o significado nutricional da digestibilidade fecal porque as determinaes so precisas, visto que o coeficiente de variao est entre 1 e 2%, e os ensaios no produzem problemas sade do animal; no obstante, recomendou manter a realizao de ambos estudos de digestibilidade para o nitrognio, visando uma melhor compreenso dos efeitos associativos que se podem apresentar entre a protena e os componentes da parede celular, principalmente no que acontece com os carboidratos que so fermentveis no intestino grosso. Por sua vez Lenis (1992) expressou que no caso dos aminocidos a situao bem diferente: por exemplo, o cido asprtico, glicina, prolina, treonina e triptofano apresentam valores de digestibilidade ileal aparente baixos e fecal altos devido a que no intestino grosso a degradao maior que a sntese. Pelo contrrio, quando neste local do trato digestivo h mais energia fermentvel, menor a quantidade de nitrognio absorvido sob a forma de amnia e, em consequncia, maior a incorporao na protena microbiana, encaminhando, portanto, a sntese de aminocidos, ocorrendo aumento de sua excreo nas fezes e decrscimo da digestibilidade entre os dois locais. Isto o que acontece com a metionina e, em proporo um pouco menor, com a lisina, tirosina, fenilalanina, isoleucina e leucina. Apesar de os estudos de digestibilidade ileal terem maiores dificuldades para sua realizao, tm vantagens sobre os de digestibilidade fecal porque as interferncias de utilizao dos aminocidos no intestino grosso so reduzidas, porm no totalmente evitadas, no tm custos muito altos e apresentam valores de desvio padro relativamente baixos (Batterham, 1992). Mas h um aspecto importante na avaliao das protenas dos alimentos em sunos, gerado pelos estudos realizados por Just et al. (1985) e confirmados por Batterham (1992), segundo os quais a digestibilidade ileal da protena e da lisina apresentou maior associao com sua disponibilidade, avaliada a partir da protena depositada, do que com a digestibilidade fecal. Partindo destes achados, nos ltimos anos tem sido sugerido utilizar os ensaios de digestibilidade ileal para estimar a disponibilidade dos aminocidos; contudo, ainda existem muito poucos dados onde possam-se comparar os resultados dos ensaios de crescimento e de digestibilidade ileal. Tm sido, ento, os princpios de digestibilidade ileal, mais que a fecal, os que serviram de base para o estabelecimento das tabelas de necessidades nos padres de alimentao para sunos na Inglaterra (ARC, 1981), nos Estados Unidos (NRC, 1988), a maioria dos pases europeos (Henry, 1985; Lenis, 1992) e no Japo (Furuya, 1994). Porm, no caso dos Estados Unidos, estes padres s consideraram, por enquanto, os valores de digestibilidade para 4 aminocidos em 18 alimentos, sem incluir ainda em suas tabelas os requisitos para sunos expressos segundo a digestibilidade ileal. Estes princpios tm sido tambm empregados nos ltimos anos por um grupo de pesquisadores da Austrlia para avaliar a utilizao e disponibilidade da lisina, treonina, metionina, triptofano, leucina, isoleucina e valina (Batterham, 1992) e esto contribuindo para o clculo dos requisitos nutricionais para aves e sunos baseados no conceito de protena ideal (Fuller & Wang, 1990; Parsons & Baker, 1994; Sve, 1994). Segundo Parsons & Baker (1994) a protena ideal refere-se condio que cabe a toda protena de possuir uma mistura de aminocidos que tenha uma composio idntica s necessidades do animal para a mantena e a mxima deposio da protena corporal,
112
sendo que estas so estimadas levando em considerao sua digestibilidade e disponibilidade biolgica. Visto que nem sempre fcil e prtico realizar estimativas das necessidades para cada aminocido, diferentes grupos de pesquisadores tm sugerido que os aminocidos indispensveis sejam expressos em relao com a lisina, considerada como o primeiro aminocido limitante das dietas usadas com freqncia na alimentao dos sunos e o segundo nas dietas para aves (Fuller & Wang, 1990; Parsons & Baker, 1994; Sve, 1994). Portanto, as estimativas das necessidades dos animais baseadas na protena ideal partem do conhecimento certo da digestibilidade e a disponibilidade da lisina nas diferentes fontes de protena. Devido s limitaes que possam surgir pela falta de dados suficientes e precisos de digestibilidade ileal aparente de aminocidos, Henry (1985) sugeriu que possvel a sua estimao multiplicando seu contedo no alimento pela digestibilidade total aparente do nitrognio. Sob estas condies, Sibbald (1987) destacou que os dados gerados so aproximados, provisrios e s teis para certos grupos de alimentos, como os farelos de soja ou de algodo. Os mtodos de digestibilidade fecal e ileal, quando comparados com as propostas in vitro, tm custos bem mais altos, so de elevada complexidade, exigem muito tempo para sua realizao e s permitem a avaliao de uma fonte de protena de cada vez. Apesar disso refletem, de forma mais exata, as relaes que existem entre o alimento e o animal, permitindo analisar todos os aminocidos de uma s vez. 3.1.2. Mtodos para estimar a disponibilidade da protena e os aminocidos dos alimentos. Como j foi dito, a digestibilidade definida como uma medida do desaparecimento da protena e dos aminocidos durante sua passagem pelo sistema digestivo e no sob a idia de absoro. Por sua vez a absoro refere-se aos intercmbios que acontecem entre o sangue e o lume digestivo. No entanto, a disponibilidade a proporo daquele aminocido limitante na rao que foi digerido, absorvido que utilizado para a sntese de protena (Batterham, 1992). Nesta parte final de reviso sobre os estudos de avaliao in vivo sero analizadas algumas das metodologias e tcnicas que existem para determinar a disponibilidade da protena e dos aminocidos dos alimentos para no ruminantes. A tabela 3 registra um resumo das mais importantes tcnicas utilizadas para avaliar de maneira integral a disponibilidade da protena nos no ruminantes. TABELA 3. TCNICAS MAIS UTILIZADAS PELOS MTODOS INTEGRAIS DE AVALIAO DA PROTENA DOS ALIMENTOS PARA NO RUMINANTES. TCNICAS Relao de Eficincia Protica (P.E.R) Relao de Protena Lquida (N.P.R) DEFINIO Ganho de peso corporal (g) Protena Bruta consumida (g) Ganho de peso corporal ajustado (g) Protena Bruta consumida (g) Ajuste: Perda de peso de um grupo
113
controle alimentado com uma rao livre de protena ndice de Crescimento de Nitrognio (I.C.N) Utilizao Bruta da Protena (B.P.U) Utilizao Lquida da Protena (N.P.U) Valor da inclinao da linha que associa o ganho de peso corporal com a ingesto de Nitrognio (b) Ganho de Nitrognio corporal (g) Nitrognio consumido (g) Ganho de Nitrognio corporal ajustado (g) Nitrognio consumido (g) Ajuste: Perda de Nitrognio de um grupo controle alimentado com uma rao livre de Protena Ganho de Nitrognio corporal ajustado (g) Nitrognio realmente digerido (g) � Ajuste: Perda de Nitrognio de um grupo controle alimentado com uma rao livre de Protena V.B = N.P.U Digestibilidade Verdadeira �
Valor Biolgico (V.B)
� A digestibilidade verdadeira estimada pelo ajuste das perdas de Nitrognio endgeno fecal dos animais controle alimentados com uma rao livre de Nitrognio. Como se pode observar na tabela anterior, existem dois tipos de propostas dentro dos mtodos de avaliao integral das protenas: a PER, NPR e ICN, que se baseam no ganho de peso corporal por unidade de protena ou nitrognio consumido e as que associam a reteno corporal de nitrognio com o nitrognio consumido (BPU e NPU) ou com o nitrognio realmente digerido (VB). Para Fuller (1988) as propostas que avaliam a protena partindo do peso corporal tm sido questionadas porque este no consiste totalmente de protena e nem sempre proporcional ao ganho de protena; alm disso, alguns resultados obtidos com a PER so afetados pelas variaes no consumo de alimento e, caso que estas no sejam controlados, possvel obter resultados errados que nem sempre dependem da qualidade da protena, mas sim da palatabilidade do alimento. Embora estes mtodos tinham sido questionados com veemncia, ainda so empregados como maneira de estabelecer anlises comparativas entre os resultados obtidos nos diferentes estudos. Segundo Fuller (1988), a relao de eficincia protica (PER), desenvolvida por Osborne & Mendel entre 1914 e 1920, o valor biolgico (V.B) de Mitchel (1924), o ndice de balano de nitrognio (I.B.N), de Allison et al. (1946) e a utilizao lquida da protena (NPU), de Bender & Miller (1953), so mtodos que tm sido empregados tradicionalmente em ratos e aves para avaliar a protena dos alimentos, o que no significa que no possam ser considerados no que diz respeito aos sunos. Para o mesmo Fuller (1988) estes mtodos fazem parte das propostas que visam avaliar as protenas de maneira integral, geram um valor nico e no aditivo para a dieta total, o
114
que no permite que seus valores sejam utilizados quando so combinadas protenas diferentes em dietas misturadas e, alm disso, no fornecem informao dos fatores que afetam o valor nutritivo. De acordo com Fuller (1988), para corregir algumas dificuldades geradas pelos mtodos de avaliao da protena que se baseam no ganho de peso corporal, em 1909 Thomas desenvolveu uma proposta na qual o valor da protena est associado eficincia de utilizao do nitrognio da dieta para promover a deposio da protena nos tecidos corporais do animal adulto. Esta proposta foi logo aperfeioada por Mitchel (1924) visando sua utilizao nos animais em crescimento para estimar a eficincia da proteina para satisfacer as necessidades de mantena dos animais, mas tambm para estimular o crescimento. O princpio deste novo mtodo, nomeado valor biolgico (VB), e algumas outras medies relacionadas com as propostas baseadas na reteno de nitrognio como so o ndice de balano de nitrognio (IBN) e a utilizao lquida da protena (NPU), aparecem registrados na figura 1, proposta por Fuller (1988). Como pode ser observado nesta fgura, na avaliao da protena se apresentam vrias relaes: Se X = Consumo de protena e Y = Ganho de peso, ento: A/I = Relao de Eficincia Protica (PER) A+B/I = Relao de Protena Lquida (NPR) Tan = ndice de Protena Lquida (IPL) Se X = Consumo de protena e Y = Reteno de Nitrognio, consequentemente: A/I = Utilizao Bruta da Protena (BPU) A+B/I = Utilizao Lquida da Protena (NPU) Se X = Consumo de protena digestvel e Y = Reteno de Nitrognio, A+B/I = Valor Biolgico (VB) Tang = ndice de Balano de Nitrognio (IBN) Segundo os protocolos de pesquisa desenvolvidos por Thomas & Mitchel o valor biolgico estimado utilizando 10% como o teor da protena na dieta; a escolha desta concentrao arbitrria, porm satisfatria pelo fato que no h mudanas importantes na forma da curva de resposta acima desta concentrao (Fuller, 1988). Por sua vez Bellaver (1994) criticou este mtodo de avaliao do valor da protena porque o nitrognio uma aproximao grosseira da real necessidade do animal, a qual expressa em termos de aminocidos e, caso que sejam estimados os valores de nitrognio endgeno, deve-se trabalhar com dietas livres de nitrognio, o que leva uma diminuio do consumo, ficando insuficiente para manter o ganho de peso, aumentando o catabolismo das protenas e superestimando as perdas de nitrognio. No que diz respeito dos aminocidos, Batterham (1992),e sua equipe de pesquisadores na Austrlia tm proposto uma metodologia, nomeada de slope-ratio, com a qual j avaliaram a disponibilidade da lisina, treonina, metionina, triptofano, leucina, isoleucina e valina, quando eles so limitantes nas dietas tpicas para sunos em crescimento. Em princpio esta metodologia expressa o grau de inclinao de uma linha de resposta das
115
dietas que contm o aminocido a ser avaliado como uma proporo do grau de inclinao da curva de resposta da dieta padro que livre do aminocido. A figura 2 registra a representao terica proposta por Batterham (1992) dos resultados obtidos em uma determinao de disponibilidade da lisina. Para estes pesquisadores, quando se planeja um experimento visando determinar a disponibilidade de um aminocido empregando esta tcnica necessrio considerar vrias condices: Segurar que a resposta animal seja devida aminocido que se est avaliando e no que esteja influenciada pelos outros nutrientes fornecidos pela fonte de protena testada. Para segurar que a resposta seja linear e corresponda com o aminocido padro, a dieta deve ser deficiente no aminocido a testar, porm adequada nos outros nutrientes; alm disso, importante incorporar a protena avaliada na dieta basal para proporcionar assm quantidades do aminocido testado que sejam semelhantes quelas que so fornecidas pela dieta padro. A adio dos outros aminocidos, alm daquele considerado limitante, provistos pela protena a avaliar, pode alterar o balano destes nas dietas, quando comparadas com a dieta padro e, consequentemente, afeta a utilizao do aminocido que est sendo avaliado. O controle na alimentao visando minimizar os efeitos dos imbalanos dos aminocidos sobre a utilizao e disponibilidade. As sugestes dos pesquisadores australianos assinalam que as dietas sero provistas baseadas no peso do animal a cada 3 horas de intervalo. Selecionar um critrio de resposta valido. Em teoria o critrio mais apropriado seria a medio da reteno corporal do aminocido avaliado, mas tambm o que tem os custos de experimentao mais altos; por consiguente, na prtica so utilizadas diferentes medidas de resposta animal para avaliar a disponibilidade do aminocido: nas aves, por exemplo, tem sido empregada a eficincia da converso alimentar porque apresenta vantagem sobre o ganho de peso devido que considera as diferenas no consumo de alimento; no entanto, nos sunos Batterham (1992) sugereu que de preferncia empregar a eficincia da converso alimentar com relao carcaa que com o peso vivo, visto que com alimentos proticos fibrosos as contribues dos nutrientes digeridos no intestino grosso dos sunos podem sobreestimar em 0,42 unidades a disponibilidade dos aminocidos. Utilizar um procedimento de anlise adequado. O anlise estatstico do mtodo slope-ratio estabelece uma regresso entre o critrio de resposta escolhido e o nvel do aminocido avaliado. Neste tipo de anlise h mecanismos estatsticos que seguram que as respostas s sejam devidas dieta e aos aminocidos estudados: Segurando que a resposta ao aminocido padro passe atravs da dieta basal, nomeada dieta branco; A suposio de linearidade da resposta animal e, consequentemente, a inexistncia de uma curva que se pode desenvolver caso que a incluso da protena avaliada produza incremento ou diminuio no consumo do alimento pela presena de um nutriente limitante na dieta basal ou um fator antinutricional. Para segurar que a resposta do aminocido avaliado seja linear necessrio realizar um experimento constitudo por 5 dietas e 4 ou 5 nveis da protena a ser avaliada.
116
Realizar um teste para segurar que as inclinaes das curvas de resposta do aminocido padro e daquele que est sendo avaliado cortem o ponto da resposta atingido com a dieta basal. A tabela 4 registra um resumo dos resultados de alguns experimentos realizados por Batterham (1992) em sunos, visando determinar a disponibilidade da lisina de vrias fontes de protena. TABELA 4. DISPONIBILIDADE DA LISINA EM FONTES PROTICAS PARA SUNOS. FONTE DE PROTENA DISPONIBILIDADE VALOR MDIO Farelo de algodo Farelo de sementes de Lupinus sp Farelo de amendoim Farelo de girasol Farinha de carne e de carne e ossos Leite em p desnatado Farelo de sementes de colza (Brassica sp) Farelo de soja Feijo (Pisum sativum) Farinha de sangue 0,34 0,55 0,57 0,60 0,68 0,85 0,87 0,88 0,93 1,08 INTERVALO 0,27-0,43 0,37-0,74 0,54-0,66 0,42-0,97 0,77-0,97 0,80-0,98 1,03-1,13
Na proposta sugerida pelos pesquisadores australianos s possvel o estudo da disponibilidade de um aminocido por vez, pelo que, em conseqncia, considerada como um mtodo que requer muito tempo, de alto custo e limitado. Alm disso, os resultados dos experimentos apresentam valores de desvio padro bem mais altos que os obtidos nos estudos de digestibilidade, devido que as estimativas baseam-se nas inclinaes de duas linhas de resposta, que por sua vez so afetadas pelas variaes individuais dos animais; mas tambm se deve ao fato que h at 10% de variao entre os laboratrios quanto determinao dos aminocidos, o qual, para Batterham (1992) afeta os ensaios de digestibilidade mais que os de crescimento: a variao de 10% na determinao dos aminocidos afeta os estimados de digestibilidade em 1%, no entanto que produz uma variao de 10% quando utilizado o crescimento animal como critrio de avaliao da disponibilidade. Embora esta proposta tinha sido criticada, Batterham (1992) assinala que os ensaios de disponibilidade so uteis para desenvolver tabelas de classificao das principais fontes de protena, pesquisar os efeitos do processamento sobre a qualidade dos alimentos e como valores padro durante o adiantamento da avaliao de novas tcnicas de
117
estimao da disponibilidade. Finalmente, os resultados dos estudos realizados por estes pesquisadores, utilizando dietas de farelo de algodo e soja, indicam que as dietas formuladas a partir da lisina total resultaram em diferenas na resposta animal (medida pelo crescimento, a eficincia da converso alimentar em relao com a carcaa, a deposio da protena e reteno da lisina) o qual no aconteceu quando as dietas foram formuladas considerando a disponibilidade da lisina. Para os mesmos pesquisadores resultaria mais apropriado, em conseqncia, a formulao de dietas baseadas nos valores de disponibilidade da lisina que na lisina total. Visto que os ensaios de disponibilidade apresentam algumas limitaes para serem empregados como mtodos rotineiros de avaliao dos aminocidos, nos ltimos anos tem sido sugerido o uso dos experimentos de digestibilidade ileal para sua estimativa, porque suponhe-se que se um aminocido digerido e absorbido no intestino delgado, ento est de forma disponvel para a sntese da protena. No obstante, que h poucos dados comparando os resultados obtidos nos ensaios de crescimento e de digestibilidade ileal, a concluso de Batterham (1992) que para as protenas de alta disponibilidade h uma relao muito prxima entre as duas variveis; no caso das protenas de baixa disponibilidade, bem por suas condies naturais ou como efeito de alguns processos de transformao, parece que a digestibilidade ileal sobreestima a disponibilidade dos aminocidos. 3. 2. O CONCEITO DE DISPONIBILIDADE NOS MTODOS DE AVALIAO in vitro. Quando se estima o valor nutricional das protenas de grande importncia a informao fornecida pelos estudos in vivo; contudo, estas metodologias requerem muito tempo e oramentos altos, razes que as converteram de utilidade limitada para serem incorporadas ao controle de qualidade rotineiro que faz a indstria produtora de raes para animais. Por estes argumentos nos ltimos anos tm surgido vrias propostas de avaliao in vitro da protena e de alguns aminocidos, as quais, de acordo com Boisen & Eggum (1991), s so vlidas se contem tcnicas simples, rpidas, reproduzveis e seus resultados estiverem associados aos valores obtidos in vivo, empregando alimentos similares e dentro de condies padronizadas. Em condies in vitro so avaliados os efeitos dos tratamentos com calor e o armazenagem dos alimentos durante longo tempo sobre a qualidade da protena e a disponibilidade dos aminocidos utilizando vrias estratgias: mtodos que utilizam a incubao com diferentes sistemas enzimticos, mtodos de solubilizao e tcnicas qumicas que estimam a disponibilidade dos aminocidos. 3.2.1. Mtodos de incubao enzimtica para avaliar a protena. Para Boisen & Eggum (1991) os mtodos in vitro que empregam incubaes devem estar acompanhados de sistemas enzimticos que tenham caratersticas semelhantes quelas presentes no trato gastrointestinal. A tabela 5 traz uma classificao dos diferentes sistemas enzimticos propostos para as incubaes in vitro. TABELA 5. CLASSIFICAO DOS SISTEMAS ENZIMTICOS EMPREGADOS NAS INCUBAES IN VITRO. SISTEMA ENZIMTICO AUTOR OBSERVAES GERAIS
118
INCUBAO DE UMA ETAPA Inculo doudenal, lquido ileal ou fezes Pepsina Tripsina Pronase Lwgren et al. (1989) Sheffner et al. (1956); AOAC (1984) Maga et al. (1973) Taverner & Farrel (1981) INCUBAO COM DUAS ETAPAS Pepsina - lquido duodenal de suno Pepsina - pancreatina Furuya et al. (1979) Clunies & Leeson (1984) Bchmann (1979) Sunos Aves Contem uma soluo de borato com caratersticas bacteriostticas O mtodo alm da pepsina e a pancreatina contem uma amilase, tem a remoo prvia dos lipdes e usa a adio de sais biliares e lipase
Babinszky et al. (1990)
INCUBAO COM TRS ETAPAS Pepsina - pancreatina lquido ruminal Pepsina - Pancreatina celulasa Verbaeke et al. (1979) Estudar a fermentao da matria orgnica no intestino grosso Contem alm lipasa, sais biliares e uma amilasa microbial (Termamyl) Estimar a digestibilidade fecal da matria orgnica De maneira geral estes mtodos tentam medir a taxa incial de hidrlise, determinar os valores mximos de digestibilidade e predizer a digestibilidade in vivo dos alimentos, processados ou no, nos no ruminantes. Dentro dos mtodos que medem a taxa incial de hidrlise da protena, com ou sem a remoo dos produtos que podem inibir a digesto, os mais promissrios so o de Hsu et al. (1977), ou de pH - drop, e o registrado por Boissen & Eggum (1991), chamado de pH -stat.
Metz & Van der Meer (1985)
119
Segundo Hsu et al. (1977), a taxa de digestibilidade de uma protena se pode calcular a partir da queda do pH aps 10 minutos de incubao da protena em uma mistura de tripsina, quimotripsina e peptidase intestinal a 37oC ou com uma incubao adicional a 55oC durante o mesmo tempo com uma proteasa de Streptomyces griseus. Quando as enzimas digerem a protena e so rompidas as ligaes dos peptdeos nas estruturas primrias, so liberados os ons H+ dos grupos carboxila livres, o que faz descer o pH da suspenso de protenas; o valor de pH quase sempre declina rapidamente no perodo inicial de incubao, estabilizando-se aos dez minutos, momento no qual medido de novo o pH. Os estudos de Hsu et al. (1977), realizados com protenas de origem vegetal, e os de Moughan et al. (1989) para vinte amostras de farinha de carne e osso registraram que houve alta correlao entre o valor de pH obtido com esta metodologia e a digestibilidade fecal aparente e ileal verdadeira determinada em ratos. Igualmente os estudos de Hsu et al. (1977) mostraram que o mtodo de pH - stat foi sensvel presena de inibidores da tripsina e a os efeitos dos tratamentos que usam calor sobre a digestibilidade da protena. Na Universidade de Illinois, Parsons (1991) encontrou que os valores obtidos com o mtodo pH - stat esteveram positiva e altamente associados com a digestibilidade in vivo da lisina de vrias fontes de protena, avaliada em aves e sunos, sendo que foram bem mais altos nas aves. Um resumo destes resultados se pode observar na tabela 6. TABELA 6. MUDANAS NOS VALORES DE pH E DIGESTIBILIDADE in vivo DA LISINA EM AVES E SUNOS DE FONTES COM ALTOS TEORES DE PROTENA. ALIMENTO MUDANAS NOS VALORES DE pH (UNIDADES) (1) 2,36 1,52 1,39 1,37 DIGESTIBILIDADE APARENTE DA LISINA (%) (2) 99 92 85 84 DIGESTIBILIDADE VERDADEIRA DA LISINA (%) (3) 86 76 78
Caseina Farelo de soja Farinha de carne Farinha de subprodutos do sacrifcio de aves Farelo de algodo Farinha de penas
(1) (2) (3)
1,23 0,90
72 68
60 -
Mtodo de pH - stat (Hsu et al., 1977) Determinada em sunos com cnula ileal Determinada com galos cecotomizados
Parsons (1991) realizou uma segunda srie de experimentos visando testar o mtodo de Hsu et al. (1977) para detectar diferenas na qualidade das protenas na farinha de penas e de carne, utilizando como critrios de avaliao os ensaios de crescimento em aves (PER e a biodisponibilidade de aminocidos) e a digestibilidade da lisina
120
determinada com galos cecotomizados. Os resultados destes estudos mostraram que no houve diferenas na digestibilidade da lisina e nos valores da PER entre as farinhas de alta e baixa qualidade, como tambm no foi encontrada associao consistente entre as mudanas nos valores de pH, a digestibilidade in vivo da lisina e a PER. O mesmo pesquisador concluiu que o mtodo de Hsu et al. (1977) sensvel para detectar diferenas na digestibilidade da lisina entre fontes proticas que variam muito na sua composio qumica, mas tem uso limitado para predizer a qualidade in vivo entre amostras diferentes da mesma fonte de protena de origem animal. Tal vez a crtica mais forte feita proposta de Hsu et al. (1997) tem a ver com o fato de que no caso das farinhas de carne e ossos possvel encontrar algumas amostras que apresentem alta digestibilidade in vivo da protena com valores baixos, no esperados, de pH, situao que explicada pela elevada capacidade tampo originada pelos altos contedos de cinzas (Moughan et al., 1989). Esta dificuldade poderia ser contornada, de acordo com Boissen & Eggum (1991), se fosse mantido constante o pH da suspenso da protena com a mistura de enzimas com a adio de uma soluo de 0,1 M de NaOH. A taxa incial de digestibilidade da protena medida, consequentemente, pelo consumo do hidrxido. Esta modalidade de avaliao da protena, quando comparada com o mtodo pH - drop, melhora a estimativa da digestibilidade fecal verdadeira das protenas de origem vegetal e animal, ainda naquelas amostras com alta capacidade tampo; contudo, por enquanto, os resultados provem de protenas com alta digestibilidade, no havendo sido registrado um nmero importante de estudos com protenas de baixa digestibilidade (Boissen & Eggum, 1991). A digestibilidade em pepsina, como mtodo enzimtico que mede os valores mximos de digestibilidade, o nico reconhecido pela Association of Official Analytical Chemits (AOAC, 1984). O principal problema desta proposta, segundo Parsons (1991), parece ser a excessiva concentrao de 0,2% de pepsina que tem sido sugerida pela AOAC (1984), visto que com ela tambm podem ser digeridas totalmente protenas de baixa qualidade. Assim, por exemplo, Johnston & Coon (1979) encontraram em nove amostras de farinhas de carne que os valores de digestibilidade para o nitrognio foram iguais quando a concentrao de pepsina caio de 0,2 para 0,02%, mas diminuieram quando esta foi reduzida a 0,002%; contudo, aumentaram de maneira importante as diferenas dos valores de digestibilidade entre as amostras a diminuir a concentrao de pepsina. Nesta mesma linha as pesquisas realizadas por Parsons (1990; 1991) levaram a resultados similares como pode ser analisado das tabelas 7 e 8 que registram um resumo de alguns dos achados obtidos por este pesquisador na Universidade de Illinois. TABELA 7. EFEITO DA CONCENTRAO DE PEPSINA SOBRE A DIGESTIBILIDADE in vitro DO NITROGNIO EM FONTES DE PROTENA DE ORIGEM ANIMAL (Valores expressos em %). CONCENT PEPSINA (%) FARINHA DE CARNE FARINHA DE PENAS FARINHA DE SUBPRODUTOS DO SACRIFCIO DE AVES MDIA 85 MNI MO 80 MXI MO 92
MDIA 0,2 86
MNI MO 83
MXI MO 89
MDIA 76
MNI MO 70
MXI MO 81
121
0,002 0,0002
72 42
54 29
83 69
32 -
17 -
49 -
75 -
58 -
89 -
Deste resultados se pode observar que os valores de digestibilidade do nitrognio diminueram quando decresceu a concentrao de pepsina, igualmente para a mesma fonte de protena aumentou a variao dos valores de digestibilidade quando foi mais baixa a concentrao de pepsina. No que tem a ver com as associaes os resultados da tabela 8 mostram que as farinhas de penas e de carne tiveram valores de correlao mais altos e estatisticamente significativos entre a digestibilidade in vitro do nitrognio com a digestibilidade da lisina quando a concentrao de pepsina foi de 0,002%, o que no aconteceu para o 0,2% de pepsina. No caso da farinha de carne, a reduo da concentrao de pepsina a 0,0002% no introduziu melhoras nos valores de correlao com a digestibilidade in vivo. Por sua vez, nas farinhas de subprodutos do sacrifcio de aves, tanto a digestibilidade do nitrognio com 0,2 como 0,002% de pepsina teveram valores de correlao altos e estatisticamente significativos com a digestibilidade da lisina. Nesta mesma linha de anlise, nas farinhas de penas e de carne os valores de correlao entre a digestibilidade in vitro do nitrognio e a disponibilidade in vivo da lisina foram mais altos e estatisticamente significativos quando utilizou-se 0,002% de pesina; mas estas mesmas amostras submetidas a 0,2% de pepsina no geraram resultados similares quanto os valores de correlao. TABELA 8. VALORES DA CORRELAO ENTRE A DIGESTIBILIDADE DO NITROGNIO EM PEPSINA COM A DIGESTIBILIDADE in vivo E A DISPONIBILIDADE DA LISINA DE PROTENAS DE ORIGEM ANIMAL. FONTE DE PROTENA NMERO DE AMOSTRAS CONCENTRAO DE PEPSINA (%) VALOR DA CORRELAO COM A DIGESTIBILIDAD E (1) 0,23 0,68 * 0,17 0,67 * 0,63 * 0,98 * VALOR DA CORRELAO COM A DISPONIBILIDAD E (2) 0,49 0,81 * 0,46 0,66 *
Farinha de penas Farinha de carne
0,2 0,002
14
0,2 0,002 0,0002
farinha de subprodutos do sacrifcio de aves
0,2
0,002
0,95 *
122
(1) A digestibilidade da lisina foi determinada em ensaios com galos submetidos a alimentao forzada. (2) A disponibilidade da lisina foi determinada utilizando o mtodo slope-ratio com frangos em crescimento. * P< 0,05 Visto que o processo de digesto muito complexo, os mtodos in vitro mais simples, que s empregam uma enzima, fornecem informao valiosa nas pesquisas que visam estudar os efeitos dos processamentos dos alimentos sobre a digestibilidade da protena (Boissen & Eggum, 1991); mas, como assinalou Parsons (1991), poderiam ser atingidos objetivos similares se fossem incorporados s avaliaes mtodos bem mais simples, como so, por exemplo, as medies de solubilidade da protena em KOH. Vrias pesquisas tm originado resultados valiosos ao serem utilizados os mtodos de duas etapas. Em sunos, por exemplo, Furuya et al. (1979) e Sakamoto et al. (1980) em experimentos realizados em aves, encontraram alta correlao entre os resultados de digestibilidade com pepsina e fluido duodenal e a digestibilidade fecal da protena e a matria seca. Contudo, ainda so pouco incorporados estes mtodos s avaliaes in vitro de disponibilidade da protena, visto as grandes dificuldades que h para padronizar as preparaes enzimticas, o que faz que seja difcil ach-las de maneira comercial. Alguns estudos relatados por Boissen & Eggum (1991) sugerem que este problema foi aprimorado de maneira certa com a substituo do lquido ileal por uma soluo pancretica, sem que sejam afetadas significativamente as relaes entre os valores in vivo e in vitro, com resultados similares aos obtidos nas pesquisas com a tcnica de saco de nilon de Sauer et al. (1989), alm que disponvel comercialmente, tem uma composio bem definida com pouca variabilidade e com resultados reproduzveis entre os laboratrios. Finalmente, no que diz respeita a este assunto, Boissen & Eggum (1991), assinalam que possvel obter valores de correlao elevados entre a digestibilidade in vivo e in vitro se so considerados trs elementos bsicos: Avaliar de forma simultnea pouco nmero de fontes de protena; Trabalhar com alimentos que tenham valores de digestibilidade com variaes relativamente altas; Estudar alimentos que pertenam a grupos prximos na sua classificao, ou com o mesmo alimento submetido a diferentes processos de transformao. De um modo geral poderia ser dito que as medies in vitro geram valores de digestibilidade verdadeira, os quais poderiam explicar porque s vezes so mais altos, quando comparados com os obtidos nos estudos in vivo de digestibilidade aparente. 3.2.2. MTODOS QUMICOS AMINOCIDOS. QUE ESTIMAM A DIOSPONIBILIDADE DOS
Como foi observado dos resultados contidos na tabela 8 a digestibilidade in vitro poderia ser usada como critrio para estimar a disponibilidade in vivo da lisina; mas acontece que para alguns alimentos processados a disponibilidade nem sempre um bom mtodo na avaliao da qualidade da protena, pelo que necessrio estudar mtodos adicionais que visem a medio in vitro da disponibilidade de alguns aminocidos, mas
123
de maneira particular da lisina porque, de acordo com Batterham (1992), o primeiro aminocido limitante das dietas para sunos e o mais susceptvel durante o processamento trmico dos alimentos. Segundo Batterham (1992), quando se aplica calor durante o processamento de um alimento os grupos -amino da lisina reagem com outros, principalmente os carbonila dos acares redutores, formando compostos de Maillard como so a fructosalisina e a lactulosalisina, os quais podem ser digeridos, porm, pouco possvel que sejam utilizados a menos que os rins tenham as enzimas necessrias para liberar a lisina, sendo, ento, excretados na urina. O desenvolvimento destas reaes reduz a digestibilidade, destri fundamentalmente a lisina, porque um aminocido dibsico, com probabilidade mexe com a treonina, metionina e triptofano, mas, por outro lado, no parece afetar os aminocidos com cadeias ramificadas, como leucina, isoleucina e valina. Visto que o teor de lisina disponvel no pode ser medido com as anlises padro que h para determinar os aminocidos devido as interferncias originadas por alguns compostos como a lactulosil - lisina, tm sido propostos vrios outros mtodos de avaliao. A tabela 9 registra as caractersticas mais importantes, vantagens e desventagens de alguns dos sugeridos por Assoumani & Nguyen (1991).
TABELA 9. MTODOS in vitro NO ENZIMTICOS PARA ESTIMAR A DISPONIBILIDADE DA LISINA NOS ALIMENTOS PARA NO RUMINANTES. MTODO VANTAGENS DESVANTAGENS QUMICOS 1-Fluor 2-4, dinitrobenzeno (FDNB) Estimao disponvel da lisina Interage com carboidratos e outros compostos Perda do DNP- Lisina na digesto cida lisina Pouco especfico
Estimao da cido2,4,6 disponvel. Tiempo trinitrobenzeno 1sulfnico Formao de ligaes colorimtricas com o uso do cido laranja
Pouco especfico Processo dficil. Sobrestima o contedo de lisina MICROBIOLGICOS (Tetrahymena pyriformis) Usado para estimar a disponibilidade in vivo ELETRODOS ENZIMTICOS Requer pretratamentos enzimticos Necessidade de coenzimas Durao das membranas
L-Lisina descarboxilase eletrodo de CO2
Alta especificidade + Pouco tempo de anlise (1-10 minutos)
124
L-Lisina oxidase eletrodo de O2
+ Especificidade Pouco tempo de anlise (1minuto)
Interfernascom ativadores no identificados Interferna com cido levulnico nas amostras hidrolizadas com cido
O primeiro mtodo qumico proposto para determinar a lisina disponvel foi desenvolvido por Carpenter & Booth (1973). Nele utiliza-se a reao dos grupos amino livres com o 1fluor-2,4- dinitrobenzeno (FDNB). Aps desta reao a amostra hidrolizada com um cido, o hidrolizado extrado com um solvente e o composto dinitrofenil - lisina (DNP lisina) que fica medido de maneira colorimtrica. O princpio desta proposta de avaliao basea-se no fato que se o grupo -amino da lisina fica livre aps o processamento da protena, ento, a lisina est disponvel e o resduo que no possui este grupo, mesmo que possa ser digervel, com toda possibilidade que no ser utilizado. Infelizmente o DNP - lisina no estvel durante a hidrlise cida da protena em presena dos carboidratos e, em conseqncia, em alguns casos possvel observar perdas de 20 a 30% (Swaisgood & Catignani, 1991). Porm a proposta de Carpenter & Booth (1973) oferea vantagens, Batterham (1992) tm criticado este mtodo por en quanto ainda no tenha sido resolvido o problema da divergncia que existe entre os valores encontrados no laboratrio e a disponibilidade in vivo de lisina, como se pode observar na tabela 10. TABELA 10. DISPONIBILIDADE DA LISINA DE FONTES PROTICAS DETERMINADA EM SUNOS EM CRESCIMENTO E COM MTODOS QUMICOS BASEADOS NO USO DO FDNB. FONTES DE PROTENA DISPONIBILIDADE QUMICA DA LISINA COM FDNB 0,97 0,90 0,90 0,84 0,83 0,80 0,80 0,65 0,79 0,77 DISPONIBILIDADE in vivo DA LISINA 1,03 0,59 0,89 0,88 0,93 0,57 0,48 0,43 0,85 0,84
Farinha de sangue Farinha de carne amostra 2 Farinha de peixe Farinha de carne amostra 3 Farelo de feijo (Pissum sativum) Farelo de amendoim Farinha de carne amostra 1 Farelo de algodo Leite em p desnatado Farelo de girasol amostra 1
125
Farelo de sementes de Lupinus sp Farelo de girasol amostra 2
0,76 0,59
0,54 0,59
Batterham (1992) afirmou que, embora no haja dvida de que o calor atue sobre a digestibilidade e a disponibilidade deste aminocido em alimentos processados, os resultados de tais associaes devem ser interpretados com precauo. Em alimentos de alta disponibilidade in vitro e para os aminocidos ramificados, esta tcnica parece til para estimar a digestibilidade ileal, porque, para eles, as mudanas na digestibilidade explicam a disponibilidade metablica. Ela menos til naqueles alimentos onde o calor diminui a disponibilidade in vitro da lisina, pois a diminuio na digestibilidade s explica uma parcela pequena da perda da disponibilidade metablica, o que, alm disso, sugere que nestes alimentos os ensaios de digestibilidade ileal no so apropriados para avaliar a disponibilidade, porque este efeito produz uma quantidade de aminocidos absorvidos em formas que so utilizadas com baixa eficincia. Existem outros mtodos qumicos para determinar a disponibilidade da lisina baseados na colorimetria, porm o mais utilizado o de FDNB porque satisfatrio visto que seus resultados so reproduzveis e permitem comparaes entre os laboratrios, alis que se mantem como o melhor procedimento para estimar a lisina disponvel nos alimentos submetidos a tratamentos com calor ou a longos perodos de armazenagem, tornandose muito mais til para qualificar ingredientes dentro do mesmo grupo de alimentos e sob condies definidas (Sibbald, 1987).
4. REFERNCIAS BIBLIOGRFICAS AGRICULTURAL RESEARCH COUNCIL. The Nutrient Requirements of Pigs. England: Commonwealth Agricultural Bureaux. p. 67-124, 1981. ARMSTRONG, G.; MOUGHAN, P.J.; MOREL, P.C.H. Comparison of methods to determine the endogenous amino acids flow at the terminal ileum of the growing rat. Journal Science Food Agriculture, v. 67, n. 3, p. 359-366, 1995. ASSOCIATION OF OFFICIAL ANALYTICAL CHEMITS. Official methods of analysis, 14th ed. Washington, DC: AOAC, 1984. ASSOUMANI, M.B.; NGUYEN, N.P. Enzyme modelling of protein digestion and Llysine availability. In: FULLER, M.F. (Ed.) In Vitro Digestion for Pigs and Poultry. Wallingford. United. Kingdom: Commonwealth Agricultural Bureaux International, p. 86-104. 1991. BABINSZKY, L. An in - vitro method for prediction of the digestible crude protein content in pig feeds. Journal Science Food and Agriculture, v. 50, p. 173-178, 1990. BATTERHAM, E.S. Availability and utilization of amino acids for growing pigs. Nutrition Research Reviews, v. 5, p. 1-18, 1992. BATTERHAM, E.S. et al. . Utilization of ileal digestible amino acids by pigs: Lysine. British Journal of Nutrition, v. 64, p. 679-690, 1990.
126
BEECH, S.A.; BATTERHAM, E. S.; ELLIOT, R. Utilization of ileal digestible amino acids by pigs: Threonine. British Journal of Nutrition, v. 65, p. 381-390, 1991. BELLAVER, C. Estimation of amino acids digestibility and its usefulness in swine feed formulation . Urbana, Champaigne: University of Illinois, 1989. 99p. (Thesis Ph. D). BELLAVER, C. Metodologias de determinao do valor das protenas e utilizao de valores disponveis nas dietas de no - ruminantes. In: Simposio Internacional de Produo de No Ruminantes. Reunio Anual da Sociedade Brasiliera de Zootecnia, 31, Maringa, Paran, 1994. Anais Maringa, Paran. Soc. Bras. Zoot., 1994. p. 1-23. BICKEL, H. III. Feed evaluation and nutritional requirements. Livestock Production Science, v. 19, n. 1-2, p. 211-216, 1988. (Special issue). BOISSEN, S.; EGGUM, B.O.Critical evaluation in vitro methods for estimating digestibility in simple-stomach animals. Nutrition Research Reviews, v. 4, p. 141162, 1991. BCHMANN, N.B. In vitro digestibility of protein from barley and other cereals. Journal Science Food and Agriculture, v. 30, p. 583-589, 1979. CARPENTER, K.J.; BOOTH, V.H. Damage to lysine in food processing: its measurement and significance. Nutrition Abstracts and Review, v. 43, n. 6, p. 423451, 1973. CLUNIES, M.; LEESON, S. In vitro estimation of dry matter and crude protein digestibility. Poultry Science, v. 63, p. 89-96, 1984. CUNNINGHAM, H.M.; FRIEND, D.W.; NICHOLSON, J.W.C. Observations on digestion in the pig using a reentrant intestinal fistula. Canadian Journal of Animal Scince, v. 43, p. 215-225, 1963. DAMMERS, J. Digestibility research in pig feeding. The influence of different factors on the digestion of feed components and the digestibility of amino acids. Leuven (Belgium), 1964. (Thesis). apud LENIS, N.P. Digestible amino acids for pigs: assessment of requirements on ileal digestible basis. Pigs News and Information, v. 13, n. 1, p. 31N-39N, 1992. DARCY, B.; LAPLACE, J.P. Digestion dans lintestine grle chez le porc. 1. Definition des conditions dobtention des digesta. Annales de Zootechnie, v. 29, n. 2, p. 137145, 1980 a. DARCY, B.; LAPLACE, J.P.; VILLIERS, P.A. Digestion dans lintestine grle chez le porc. 2. Cintique compare de passage des digesta selon le mode de fistulation, ilo-caecale ou ilo-colique post-valvulaire, dans diverses conditions dalimentation. Annales de Zootechnie, v. 29, n. 2, p. 147-177, 1980 b. DARCY, B.; LAPLACE, J.P.; VILLIERS, P.A. Obtention des digesta parvenant au gros intestine par fistulation ilo-colique post-valvulaire: Note prliminaire. Reprodution Nutrition Dvelopment, v. 20, n. 4B, p. 1197-1202, 1980 c. DARCY-VRILLON, B.; RRAT, A. Protein digestion and absorption of the hydrolysis product in the small intestine of the pig. Colloques de lI.N.R.A, v. 1, n. 16, p. 233244, 1983. apud. HENRY, Y. Principles of protein evaluation in pig feeding. World Review of Animal Production, v. 21, n. 4, p.47-59, 1985.
127
DARCY-VRILON, B.; LAPACE, J.P. Digestibilitie ilale: Utilisation compare de deux mthodes de collecte dans le cas de rgimes riches en fibre. Reprodution Nutrition Dvelopment, v. 25, n.4B, p. 847-848, 1985 a. DARCY-VRILLON, B.; LAPLACE, J.P. Ileal amino acid digestibility measurement in pigs fed high fiber diets: ileo-rectal anastomosis versus ileo-colic post-valve fistulation. In: Digestive Physiology in the Pig. Proceeding 3rd International Seminar. Copenhagen, Denmark, 16th-18th, 1985. JUST, A.; JORGENSEN, H.; FERNANDEZ, J.A. National Institute of Animal Science, p. 184-187 b. DARCY-VRILLON, B.; LAPLACE, J.P. Digesta collection procedure may affect ileal digestibility in pigs fed diets based on wheat bran or beet pulp. Animal Feed Science and Technology, v. 27, n. 4, p. 307-316, 1990. De LANGE, C.F.M. et al. The effect of feeding different protein-free diets on the recovery amino acids composition of endogenous protein collectes from the distal ileum and feces in pigs. Journal Animal Science, v. 67, p. 746-754, 1989. DECUYPERE, J.A. et al. Gastrointestinal cannulation in pigs: A simple technique allowing multiple replacements. Journal of Animal Science, v. 46, n. 3, p. 463-468, 1977. DONKOH, A.; MOUGHAN, P.J.; SMITH, W.C. Comparison of the slaughter method and simple T-piece cannulation of the terminal ileum for determining ileal amino acid digestibility in meat and bone meal for the growing pig. Animal Feed Science and Technology, v. 49, p. 43-56, 1994 a. DONKOH, A.; MOUGHAN, P.J.; SMITH, W.C. The laboratory rat as a model for determining ileal amino acid digestibility in meat and bone meal for the growing pig. Animal Feed Science and Technology, v. 49, p. 57-71, 1994 b. DONKOH, A.; MOUGHAN, P.J.; SMITH, W.C. True digestibility of amino acids in meat and bone meal for the growing pig-application of a routine rat digestibility assay. Animal Feed Science and Technology, v. 49, p. 73-86, 1994 c. DONKOH, A.; MOUGHAN, P.J.; MOREL, P.C.H. Comparison of methods to determine the endogenous amino acid flow at the terminal ileum of growing rat., JournalScience Food and Agriculture, v. 67, p. 359-366, 1995. DOURMAD, J-Y.; LE MOUL, C.; RAINELLI, P. Rduction des rejets azots des porcs par la voie alimentatione: valuation conomique et influence des changements de la Ploitique Agricole Commune. Productions Animals, v. 8, n. 2, p. 135-144, 1995. EASTER, R. A.; TANKSLEY, T.D. A technique for re-entrant ileo cecal cannulation of swine. Journal of Animal Science, v. 36, n. 6, p. 1099-1103, 1973. EGGUM, B.O A study of certain factors influencing protein digestibility in rats and pigs. Copenhagen, 1973. (Thesis). apud LENIS, N.P. Digestible amino acids for pigs: assessment of requirements on ileal digestible basis. Pigs News and Information, v. 13, n. 1, p. 31N-39N, 1992. FLATT, W.P. Feed Evaluation Systems: Historical Background. p. 1-2; 4. In: ORSKOV, E.R. World Animal Scince. B. Disciplinary Approach. 4. Feed Science. Elsevier Scince Publishers. B.V. 1988. FULLER, M.F. Methods of protein evaluation for non ruminantes. p. 81-101. In: ORSKOV, E.R. World Animal Science. B. Disciplinary Approach. 4. Feed Science. Elsevier Publishers. B.V. 1988.
128
FULLER, M.F.; WANG, T.C. Digestible ideal protein- a measure of dietary protein value. Pigs News and Information, v. 11, n. 3, p. 353-357, 1990. FULLER, M.F. et al. The mesurement of dietary amino acid digestibility in pigs, rats and chickens: A comparison of methodologies. Animal Feed Science and Technology, v. 48, p. 305-324, 1994. FURUYA, S. An outline of the japanese feeding standard for swine revised in 1993. Pigs News and Information, v. 15, n. 3, p. 81N-85N, 1994. FURUYA, S.; KAJI, Y. Estimation of the true ileal digestibility of amino acids and nitrogen from their apparent values for growing pigs. Animal Feed Science and Technology, v. 26, p. 271-285, 1989. FURUYA, S. et al. A new in vitro method for the estimation of digestibility using the intestinal fluid of the pig. British Journal of Nutrition, v. 41, p. 511-520, 1979. GARGALLO, J.; ZIMMERMAN, D.R. A simple intestinal cannula for swine. American Journal Veterinary Research, v. 41, n. 4, p. 618-619, 1980. GIDENNE, T. et al. Effect of ileal cannulation on rabbit digestion and caecotrophy: An interlaboratory study. World Rabbit Science, v. 2, n. 3, p. 101-106, 1994. GIRALDO, M.AM. Mtodos de colheita do contedo ileal para estimar a digestibilidade da protena e dos aminocidos em sunos. Belo Horizonte, Minas Gerais: Universidade Federal de Minas Gerais, 1996. 8 p. Seminrio de Zootecnia, Escola de Veterinria. HENRY, Y. Principles of protein evaluation in pig feeding. World Review of Animal Production, v. 21, n. 4, p.47-49, 1985. HSU, H.U. et al. A multienzymatic technique for estimating protein digestibility. Journal of Food Science, v. 42, p. 1269-1273, 1977. JOHNSON, R.J. Principles, problems and application of amino acid digestibility in poultry. Worlds Poultry Science Journal, v. 48, n. 3, p. 232-246, 1992. JOHNSTON, J.; COON, C.N. The use of varying levels of pepsin for pepsin digestion studies with animal proteins. Poultry Scince, v. 58, p. 1271-1273, 1979. JONDREVILLE, C. Availability of amino acids in feestuffs for pigs. Feed Mix, v. 2, n. 4, p. 10-12, 1994. JUST, A.; JORGENSEN, H.; FERNNDEZ, J.A. The digestive capacity of the caecumcolon and the value of the nitrogen absorved from th ehind gut for protein syntesis in pigs. British Journal of Nutrition, v. 46, p. 209-219, 1981. JUST, A.; JORGENSEN, H.; FERNNDEZ, J.A. Correlations of protein deposited in growing female pigs to ileal and faecal digestible crude protein and amino acids. Livestock Production Science, v. 12, p. 145-159, 1985. KELEN, C.J. van der. et al. Sources of variation of the in situ nylon bag technique. Animal Feed Science and Technology, v. 38, p. 35-42, 1992. KLHER, T. et al. Effect of ileo-rectal anastomosis and post-valve T-caecum cannulation on growing pigs. 1. Growth performance, N-balance and intestinal adaption. British Journal of Nutrition, v. 68, p. 293-303, 1992. KUIKEN, K.A.; LYMAN, C.M. Availability of amino acids in some foods. Journal of Nutrition, v. 36, n. 3, p. 359-368, 1948.
129
LAPLACE, J.P. et al. Measurement of precaecal dietary protein and plant cell wall digestion in pigs; comparison of four surgical procedures for ileo-rectal anastomosis. Livestock Production Science, v. 40, n. 3, p. 313-328, 1994. LEEUWEN. P. van. et al. Methodolical aspects for determination of amino acid digestibilities in pigs fitted with ileocecal re-entrant cannulas. Journal Animal Physilogy and Animal Nutrition, v. 58, p. 122-133, 1987. LEEUWEN, P. van, et al. A new technique for collection of ileal chyme in pigs. In: Digestive Physiology in the Pig. Proceeding 4th International Seminar. Jablona, Polish, 1988. BURACZEWSKA, L.; PASTUSZEWSKA, S.; ZEBROWSKA,T. Polish Academic of Sciences, p. 289-296. LEEUWEN, P, van. et al The post valve T-cecum cannulation technique in pigs applicated to determine the digestibility of amino acid in maize, groundnut and sunflower meal. Journal Animal Physiology and Animal Nutrition, v. 65, n. 3-4, p. 183-193, 1991. LEIBHOLZ, J. A rapid assay for the measurement of protein digestion to the ileum of pigs by the use of a mobile nylon bag technique. Animal Feed Science and Technology, v. 33, p. 209-219, 1991. LENIS, N.P. Digestible amino acids for pigs: assessment of requirements on ileal digestible basis. Pigs News and Information, v. 13, n. 1, p. 31N-39N, 1992. LOW, A.G. Nutrient absorption in pigs. Journal Science Food and Agriculture, v. 31, p. 1087-1130, 1980. LOW, A.G. Digestibility and availability of amino acids from feedstuffs for pigs: a review. Livestock Production Scince, v. 9, p. 522-520, 1982. LWGREN, W.; GRAHAM, H.; AMAN, P. An in vitro method for studying digestion in the pig. 1. Simulating digestion in the different compartments of the intestine. British Journal of Nutrition, v. 61, p. 673-687, 1989. MAGA, J.A.; LORENZ, K.; ONAYEMI, O. Digestive acceptability of proteins as measured by the initial rate of in vitro proteolysis. Journal of Food Science, v. 38, p. 173-174, 1973. MASON, V.C. Metabolism of nitrogenous compounds in the large gut. Proceedings of the Nutrition Society, v. 43, p. 45-53, 1984. METZ, S.H.M.; VAN der MEER, J.M. Nylon bag and in vitro techniques to predict in vivo digestibility of organic matter in feedstuffs for pigs In: Digestive Physiology in the Pig. Proceeding 3rd International Seminar. Copenhagen, Denmark, 16th-18th, 1985. JUST, A.; JORGENSEN, H.; FERNANDEZ, J.A. National Institute of Animal Science, p. 373-376. MITCHEL, H.H. The nutritive value of proteins. Physiological Reviews, v. 4, p. 424-478, 1924. MITCHEL, H.H. Comparative nutrition of man and domestic animals. New York: Academic Press. v. 2, 612p, 1964. apud. SIBBALD, I.R. Estimation of bioavailable amino acids in feedstuffs for poultry and pigs: a review with emphasis on balance experiments. Canadian Journal of Animal Science, v. 76, n. 2, p. 221-300, 1987. MOUGHAN, P.J.; SMITH, W.C.; JAMES, K.A.C. Preliminary observations on the use of the rat as a model for the pig in the determination of apparent digestibility of
130
differents proteins. New Zealand Journal Agriculture Research, v. 27, n. 4, p. 509512, 1984. MOUGHAN. et al. In - vitro determination of nitrogen digestibility and lysine availability in meat and bone meals and comparison with in - vivo ileal digestibility estimates. Journal Science Food and Agriculture, v. 47, p. 281-292, 1989. MOUGHAN, P.J. et al. Assessment of apparent ileal lysine digestibility for use in diet formulation for the growing pig. Animal Feed Science and Technology, v. 34, p. 95109, 1991. MOUGHAN, P.J. et al. Evaluation of the isotope dilution technique for determining ileal endogenous nitrogen excretion in the rat. Journal Science Food and Agriculture, v. 58, n. 2, p. 165-172, 1992. NATIONAL RESEARCH COUNCIL. Nutrient Requirements of Swine. 9. ed. Washington, D.C: National Academic Science, 1988. 93p. NECESIDADES nutritivas de los animales domsticos. Leon, Espaa: Editorial Academia. n 3. Cerdos, 324p, 1978. OSLAGE, H.G. et al. The methodical problems involved in investigating the intestinal breakdown of plant cell wall constituents in pigs. Animal Research and Development, v. 29, p. 111-12o, 1989. OLSZEWSKI, A. Absorption of amino acids in the cecum of pigs. Jablona (Polonia), 1975. (Ph. D. Thesis). apud. SAUER, W.C.; OZIMEK, L. Digestibility of amino acids in swine: results and their practical applications. A review. Livestock Production Science, v. 15, p. 367-388, 1986. PAPADOPOULOS, M.C. Estimation of amino acid digestibility and availability in feedstuffs for poultry. Worlds Poultry Science Journal, v. 41, n. 1, p. 64-71, 1985. PARSONS, C.M. Digestibility of amino acids in feedstuffs for poultry. In: Proceedings Nutrition Conference for Feed Manufactures. March 22-23, 1990. p. 22-29. Departments of Poultry and Animal Sciences, University of Mariland. PARSONS, C.M. Use of pepsin digestibility, multyenzime pH change and protein solubility assays to predict in vivo protein quality of feedstuffs. In: FULLER, M.F. (Ed.) In Vitro Digestion for Pigs and Poultry. Wallingford. United. Kingdom: Commonwealth Agricultural Bureaux International, p. 105-115. 1991. PARSONS, C.M.; BAKER, D.H. The concept and use of ideal proteins in the feeding of non ruminants. In: Simposio Internacional de Produo de No Ruminantes. Reunio Anual da Sociedade Brasiliera de Zootecnia, 31, Maringa, Paran, 1994. Anais Maringa, Paran. Soc. Bras. Zoot., 1994. p. 119-128. RAHARJO, Y.; FARREL, D.J. A new biological method for determining amino acid digestibility in poultry feedstuffs using a simple cannula, and the influence of dietary fibre on endogenous amino acid output. Animal Feed Science and Technology, v. 12, p. 29-45, 1984/1985. RRAT, A. Absorption of nitrogen and amino acids from exogenous (fish meal proteins) or endogenous sources in the pig. Pigs News and Information, v. 11, n. 2, p.173-180, 1990.
131
ROSTAGNO, H.S.; PUPA, J.M.R.; PACK, M. Diet formulation for broilers based on total versus digestible amino acids. Journal Applied Poultry Research, v.4, p.1-7, 1995. ROWAN, A.M.; MOUGHAN, P.J..; WILSON, M.N. The flows of deoxyribonucleic acid and diaminopimelic acid and the digestibility of dietary fibre components at the terminal ileum, as indicators of microbiano activity in the upper digestive tract of ileostomised pigs. Animal Feed Science and Technology, v.36, p. 129-141, 1992. SAHA, D.C.; GILBREATH, R.H. A modified chromic oxide indicator ratio technique for accurate determination of nutrient digestibility. Canadian Journal Animal Science, v. 73, n. 4, p. 1001-1004, 1993. SAKAMOTO, K. et al. Estimation of in vivo digestibility with laying hen by an in vitro method using the intestinal fluid of the pig. British Journal of Nutrition, v. 43, p. 389391, 1980. SAUER, W.C.; OZIMEK, L. Digestibility of amino acids in swine: results and their practical applications. A review. Livestock Production Science, v. 15, p. 367-388, 1986. SAUER, W.C. et al The evaluation of the mobile nylon bag technique for determining the apparent protein digestibility in a wide variety of feedstuffs for pigs. Journal of Animal Scince, v. 67, n. 2, p.432-440, 1989. SVE, B. Alimentation du porc en croissance: intgration des concepts de protine idal, de disponibilt digestive des acides amins et denergie nette. Productions Animales, v. 7, n. 4, p. 275-291, 1994. SHEFFNER, A.L.; ECCKFELDT, G.A.; SPECTOR, H. The pepsin - digest - residue (PDR) amino acid index of net protein utilization. Journal of Nutrition, v. 60, p. 105120, 1956. SIBBALD, I.R. Estimation of bioavailable amino acids in feedingstuffs for poultry and pigs: a review with emphasis on balance experiments. Canadian Journal of Animal Science, v. 67, n. 2, p. 221-300, 1987. SKILTON, G.A.; SMITH, W.C.; MOUGHAN, P.J. The ileal digestibility of nitrogen and amino acids in meat and bone meals determined using a rat assay. Animal Feed Science and Technology , v. 34 n. 1-2, p. 111-126, 1991. SWAISGOOD, H.E.; CATIGNANI, G.L. Protein digestibility: In vitro methods of assessment. Advances in Food and Nutrition Research, v. 35, p. 185-236, 1991. TAVERNER, M.R.; FARREL, D.J. Availability to pigs of amino acids in cereal grains. 3. A comparison of ileal availability values with fecal, chemical and enzymic estimates. British Journal of Nutrition, v. 46, p. 173-180, 1981. THOMAS, P.C. Feed evaluation- Energy. p. 66-67. In: ORSKOV, E.R. World Animal Science. B. Disciplinary Approach. 4. Feed Science. Elsevier Science Publishers. B.V. 1988. THORPE, E.; THOMLINSON, J.R. Autolysis and post-mortem bacteriological changes in the alimentary tract of the pigs. Journal Pathology Bacteriology, v. 67, p. 601-610, 1967.
132
VERVAEKE, I.J. et al. Quantitative in vitro evaluation of the energy metabolism influenced by virginiamycin and spiramycin used as growth promotors in pig nutrition. Journal of Animal Science, v. 49, p. 846-856, 1979. WALKER, W.R.; MORGAN, G.L.; MAXWELL, C.V. Ileal cannulation in baby pigs with a simple T-cannula. Journal of Animal Scince, v. 62, p. 407-411, 1986. WILLIAMS, P.E.V. Digestible amino acids for non-ruminant animals: theory and recent challenges. Animal Feed Science and Technology, v. 53, p. 173-187, 1995. ZEBROWSKA, T. Digestion and absorption of nitrogenous compounds in the large intestine of pigs. Rocz. Nauk. Rol, B-95: 85, 1973. apud. ZEBROWSKA, T. Determination of available amino acids in feedstuffs for monogastris. Feedstuffs, v. 50, n. 53, p. 15-17; 43-44, 1978. ZEBROWSKA, T. Determination of available amino acids in feedstuffs for monogastrics. Feedstuffs, v. 50, n. 53, p.15-17; 43-44, 1978.
METODOLOGIA DE AVALIAO ENERGTICA DOS ALIMENTOS PARA NO RUMINANTES I. INTRODUO A energia vital para todos os seres. Em ltima anlise toda energia utilizada pelos seres vivos provm do sol, j que as plantas captam a energia solar e armazenam essa energia em compostos qumicos. Os animais, ingerindo as plantas, iro desdobrar esses compostos para obterem a energia de que necessitam e que ser armazenada nas formas de carboidratos, gorduras e protenas. Sabe-se que a maior parte do alimento consumido por um animal qualquer utilizada para a produo de energia. Dentre os constituintes dos alimentos, os carboidratos, as gorduras, os leos e as protenas so os grandes fornecedores de energia para o organismo animal. As vitaminas e outras substncias tambm podem fornec-la, em quantidades pequenas. A porcentagem de energia total que pode ser retida pelo organismo e utilizada para realizar os processos metablicos dependem da habilidade do animal para digerir o alimento. Pelo processo de digesto e mediante a degradao dos compostos qumicos complexos em molculas de mais fcil absoro, se d a ingesto de energia. A energia total do alimento ingerido pelo animal, no totalmente aproveitada existindo perdas, razo pela qual a energia de um alimento pode ser expressa de diversas maneiras, cada uma correspondendo a um valor energtico em termos de calorias conforme o alimento utilizado (Fig. 1). A avaliao dos alimentos um dos pontos bsicos mais importantes para uma boa nutrio. atravs desta anlise que conseguiremos informaes bsicas relacionadas aos alimentos e nutrientes. Um dos mtodos mais antigos de avaliao dos alimentos o mtodo das anlises proximais ou mtodo de Weende, desenvolvido em 1864, na Alemanha. Este um mtodo de anlise qumica simplificado, rpido e barato, que ainda usado atualmente para avaliao da maioria dos alimentos, objetivando a formulao de raes. Atravs deste mtodo possvel
133
determinar: matria seca, protena, extrato etreo, extrato no nitrogenado, fibra bruta e cinzas. A energia de um alimento, bem como as necessidades energticas dos animais, so medidas em calorias. Em nutrio animal a medida de aferio a grande caloria ou quilocaloria (kcal) e significa a quantidade de calor necessria para elevar de 1C a massa de 1 kg de gua na temperatura de 14,5C. Correspondente quilocaloria temos a pequena caloria ou caloria (cal) e a megacaloria (Mcal) que definem a quantidade de calor necessria para elevar de 14,5 15,5C as massas de 1 grama e 1 tonelada de gua, respectivamente (Andriguetto et al., 1982). As unidades utilizadas na medio da energia consumida (dos alimentos) ou produzida pelo animal so expressas em termos de concentrao de energia por unidade de peso: caloria por grama (cal/g), kcal / g, kcal / kg, Mcal / kg. O joule (J) tem sido usado, principalmente na Europa, onde a unidade oficial (1,0 J = 0,239 cal ou 1,0 cal =4,18 J). Diversas determinaes realizadas sobre glicdeos e lipdeos e protenas, mostraram os seguintes valores energticos mdios por grama de matria seca: Glicdeos -------------------------- 4,15 cal Lpideos --------------------------- 9,40 cal Protenas --------------------------- 5,65 cal Os cidos graxos volteis (AGV) ou de cadeia curta so produzidos em grande quantidade na digesto dos animais ruminantes atravs da fermentao, sendo sua principal fonte primria de energia. Contudo, so tambm produzidos em menor proporo no trato digestivo de humanos e outros animais onvoros como por exemplo os sunos. Estas estimativas de produo de AGV no ceco e intestino delgado, so difceis pela inacessibilidade destes locais e tm sido obtidas atravs de vrios mtodos como por exemplo: diluio isotpica, incubao in vitro, e atravs da tcnica da canulao ou medio sangnea do contedo de AGV. Em geral, as estimativas so de que a contribuio dos AGV para complementar os requerimentos energticos dos animais so: Ruminantes ---------------- 70% Humanos ------------------- 10% Outros onvoros ---------- 20-30% Geralmente, a qualidade da dieta influi na produo de AGV, exemplo: aumento de fibra. Alm disso, existem indicaes de que os AGV exercem influncia no metabolismo de secreo da insulina, glucagon e tambm no prprio metabolismo do colesterol sendo importante ento, o desenvolvimento de um mtodo preciso para mensurao da produo dos AGV nos animais no ruminantes. O objetivo desta reviso o de avaliar criticamente os mtodos utilizados para mensurar o nvel de energia contido nos alimentos utilizados para nutrio de monogstricos.
134
Figura 1: Distribuio e utilizao da energia consumida pelos animais. Energia bruta (Energia do alimento)
Energia fecal
Energia digestvel
Energia da urina
Energia metabolizvel
Energia usada no metabolismo dos nutrientes Energia lquida
Energia usada para: - Mantena - Metabolismo basal - Atividade voluntria - Temperatura corporal
Energia produtiva - Crescimento - Reproduo - Gestao - Lactao.
135
II. UNIDADES PRTICAS DE ENERGIA: 1. Energia bruta (EB): A energia dos alimentos qumica ou potencial, ou seja, a energia que une os tomos das molculas orgnicas. Essa energia no medida diretamente. A estimativa dos nveis de energia feita a partir da oxidao total do alimento em oxignio puro uma presso de 30 atmosferas de oxignio. Esta energia desprendida da queima total do alimento denominada energia bruta (EB), pois no h qualquer indicao de quanto o animal pode dela aproveitar, ou mesmo se pode aproveit-la. Este o ponto de partida para a determinao de toda a energia utilizada pelos animais e para a avaliao nutricional dos alimentos. A energia bruta sofre partio dentro do organismo. Sinnimo: calor de combusto. Energia bruta consumida (Ebc) a energia bruta do alimento consumido multiplicado pela EB do alimento por unidade de peso seco. Ebc = EB do alimento x MS do alimento
2. Energia digestvel (ED): a energia bruta (EB) do alimento consumido menos a energia das fezes (EF), que por sua vez ser maior quanto menor for o coeficiente de digestibilidade da matria seca do alimento ou de seus princpios nutritivos. Mais apropriada para ser utilizada como unidade de alimentos utilizados para monogstricos, pois nos ruminantes, ainda temos a energia dos gases combustveis e o prprio calor desprendido durante a fermentao, o que incorreria em erros. ED = EB - EF As fezes so compostas pela poro no digerida e no absorvida do alimento, mais microorganismos e descamaes, enzimas e muco do trato gastrointestinal, principalmente. Este material d um valor energtico s fezes, valor este que se perde. Por outro lado, boa parte do contedo das fezes no provm do alimento, mas do prprio organismo animal, de origem endgena, e no deveria ser computado, pois mascara a verdadeira digestibilidade da energia do alimento. Quando esta frao endgena medida tem-se a energia digestvel verdadeira. Na maioria das vezes, o que se tem energia disgestvel aparente.
3. Energia metabolizvel (EM) : um valor bem mais seguro, pois leva em conta a energia desprendida nas fezes (EF), na urina (EU) e nos gases (EG), dando portanto, uma melhor estimativa do que estaria ocorrendo.
136
EM = EB - EF - EU - EG
A energia da urina composta de uma frao no utilizada dos nutrientes absorvidos (perda varivel) e outra, endgena, proveniente do prprio organismo animal (perda fixa). A perda urinria fixa, encontrada por Atwater ao estudar o valor da EM de alimentos para seres humanos, de 1,25 cal/g de protena, em virtude da energia potencial da uria excretada. A energia bruta dos gases medida por meio de calormetros especiais, que recolhem e medem a quantidade excretada na respirao ou pelo nus. Dentre os gases produzidos no trato gastrointestinal e resultantes das fermentaes microbianas o mais importante o metano (CH4), produzido em grande quantidade pelos ruminantes. Outros gases produzidos em menores quantidades so: hidrognio, monxido de carbono, acetona, etano e sulfeto de hidrognio. Nos monogstricos a quantidade de metano pequena e, por isso, geralmente omitida nos clculos da energia metabolizvel. Por isto, a perda de energia pelos gases somente tem significncia nos ruminantes. 3.1. Energia metabolizvel corrigida para Nitrognio (EMc): a energia metabolizvel total corrigida do nitrognio retido ou perdido pelo corpo. O balano de nitrognio (BN) a diferena entre o nitrognio consumido (Nc) e o nitrognio eliminado (Ne ) nas fezes e urina: BN = Nc - ( N fezes + N urina) Quando a diferena for igual a zero o animal est em equilbrio. Porm, em trabalhos de preciso ns teremos outros tipos de perdas como: escamao da pele, perdas por expirao, perdas de produo (leite, ovos, etc...). Quando o animal no se apresenta em equilbrio (balano nitrogenado igual zero) e o balano de nitrognio for positivo ou negativo devemos aplicar um fator de correo para o clculo da energia metabolizvel. Para mamferos a correo feita da seguinte maneira: - para cada grama de nitrognio urinrio derivado do catabolismo das protenas do corpo (igual ao balano negativo do nitrognio), 7,45 kcal o valor a ser adicionado EM, - para cada grama de nitrognio retido no corpo (igual ao balano positivo de nitrognio), o valor 7,45 ser subtrado da EM. EMc = EB - EF - EU - EG (BN x 7,45) Este valor foi obtido com ces no sendo, possivelmente, inteiramente correto para outros animais. Para animais produtores de leite ou ovos, no feita a correo para o nitrognio desses produtos. Em aves, o fator comumente usado 8,22 kcal, porque esse valor representa o equivalente energtico do cido rico por grama de nitrognio. Algumas vezes usado o fator 8,7 kcal, porque representa o contedo mdio de energia da urina por grama de nitrognio. A energia metabolizvel tambm pode ser calculada em funo da energia digestvel, atravs da seguinte frmula:
137
EM = 0,82 . ED 4. Energia lquida (EL): Energia lquida a energia metabolizvel menos o incremento calrico (IC), ou seja, a energia efetivamente utilizada pelo organismo, para sua prpria manuteno ou para produo. EL = EM - IC Incremento calrico (IC) o aumento da produo de calor observada no organismo animal aps a ingesto de alimentos, quando o animal estiver em um meio termicamente neutro. A zona de neutralidade trmica a faixa de temperatura na qual o animal pode manter a sua temperatura estvel, apenas acionando os mecanismos fsicos de regulao de temperatura. Quando o animal est em um ambiente com temperaturas mais baixas que a temperatura crtica a energia do incremento calrico ser usada para manter o corpo aquecido e assim torna-se parte da energia lquida exigida para mantena. O incremento calrico composto na prtica por duas fraes de calor: - Calor produzido pela fermentao do tubo digestivo. - Calor do metabolismo celular. Um mtodo consistente na mensurao do IC o de subtrair a produo de calor do animal em jejum, pela produo de calor do animal quando alimentado. Se no for possvel manter o animal em jejum, a produo de calor poder ser determinada alimentando-o com 2 ou mais nveis de consumo de nutrientes e calculando a diferena na produo de calor. Os nveis de alimentos devem estar prximos aos das funes fisiolgicas para os quais os dados devem ser aplicados (Harris, 1970).
IC = Produo de calor de um animal em jejum - Produo de calor em um animal alimentado A produo do incremento calrico, varia at com a natureza do alimento. Alimentos concentrados apresentam menor incremento calrico do que fibras. Os dados obtidos de energia lquida, no so muito confiveis, da no ser usada com freqncia nos clculos de rao. Os erros esto muito relacionados temperatura ambiental. Um dado obtido em uma zona fria nunca se eqivaler a um dado obtido em uma zona quente (tropical), pois quando a temperatura ambiente inferior zona neutra o animal ter que acionar mecanismos qumicos para manter a temperatura corporal constante. Quando se usa EL, deve-se estabelecer claramente quais as funes includas. Por exemplo: - EL para mantena: a frao da EL total gasta para manter o animal em equilbrio energtico. Neste estado no h ganho ou perda de energia lquida nos tecidos corporais. A EL para mantena calculada pela soma do metabolismo do animal em jejum , atividades voluntrias do organismo e energia gasta com a homeostase. O metabolismo em jejum obtido em ambiente de neutralidade trmica, no estado psabsortivo, o animal deve estar consciente e em completo repouso.
138
- EL de produo: a frao da EL necessria para o trabalho involuntrio, para o ganho em tecido (crescimento e gordura), ou para a sntese do feto, leite, ovos, l, pele, penas, etc... 6. Energia produtiva (EP): a frao da energia que realmente ser utilizada para a produo: lactao, produo de ovos, reproduo, ganho de peso ou crescimento, deposio de gordura ou de protena, etc... A energia digestvel e metabolizvel so utilizadas em ruminante, sunos, eqinos, coelhos e outros mamferos. No caso das aves, a separao de fezes e urina complicada, e somente usada a energia metabolizvel. A utilizao mdia da energia dos alimentos em sunos em fase de crescimento, considerando-se a EB de um alimento como 100%, seria de 82% de ED, 78,7% de EM e 58,2% de EL. V. MTODOS DE AVALIAO ENERGTICA DOS ALIMENTOS 1. MTODOS QUMICOS: 1.1. NDT: Este mtodo de se avaliar a energia de um alimento parece ter sua origem na Estao Agrcola de Vermont, datando de 1900. O NDT, que foi usado por muitos anos, tem sido gradualmente substitudo por outros mtodos de anlises de energia e atualmente tem seu uso restrito praticamente para bovinos, e em alguns casos, para sunos. Os valores de nutrientes digestveis totais (NDT) nos permitem comparar a concentrao de energia digestvel entre os ingredientes da dieta. Sabe-se que unicamente para as protenas, no que diz respeito energia metabolizvel, h o decrscimo de 1,25 calorias por grama. Considerando esta perda de 1,25 calorias por grama de protena absorvida vamos verificar que temos a seguinte relao calrica, por grama de nutrientes: Protena bruta digestvel ---------------------- 5,65 - 1,25 = 4,4 cal 4,0 Fibra bruta digestvel -------------------------- 4,15 cal 4,0 Extrativos no nitrogenados digestveis -- 4,15 cal 4,0 Extrato etreo digestvel ---------------------- 9,4 cal 9,0 Observamos portanto, que existe uma relao de 4:9 entre protena digestvel e glicdios digestveis respectivamente, relao que pode ser simplificada para 1:2,25. Isto quer dizer que na frao digestvel dos componentes orgnicos dos alimentos, os lipdios apresentam um valor energtico 2,25 vezes maior que os demais. Portanto, para o clculo de NDT o extrato etreo multiplicado por 2,25 para ajustar a maior concentrao de energia da gordura. Partindo deste raciocnio, aplica-se a seguinte frmula: PB digestvel + FB digestvel + ENN digestvel + (EE digestvel x 2,25) = Equivalncia energtica dos alimentos (%).
139
A energia digestvel pode ser calculada partir dos nutrientes digestveis totais usando-se a seguinte frmula: ED = % NDT : 100 x 4,405 O NDT tem sido criticado porque ele d protena o mesmo peso que os carboidratos, e tambm porque uma soma de vrias anlises de digestibilidade. Portanto, o erro associado com cada anlise individual ir reduzir a acurcia deste valor. Alm disto no considera a perda dos gases por eructao nos ruminante, bem como no considera o incremento calrico. Em face da influncia que o teor em fibra bruta exerce na digestibilidade dos alimentos, provvel que a mistura de alimentos, em uma rao, tenha seu valor em NDT ligeiramente alterado, tanto mais quanto maior for a incluso de alimentos fibrosos (Andriguetto et al, 1982). Outros pontos negativos deste sistema so: - no mede a energia em unidades energticas, expressando-a em percentagem. - est baseado em valores de combusto fisiolgica no homem e ces, no sendo aplicveis aos ruminantes (Harris, 1970). 1.2. SISTEMA CALORIA Em novembro de 1958 o Comit de Nutrio Animal do Conselho Nacional de Pesquisas dos EUA tomou a resoluo de iniciar o uso do sistema da caloria, juntamente com o NDT, para descrever o valor energticos dos alimentos, raes e exigncias nutricionais dos animais. Energia bruta: Uma amostra do material a ser testado pesada na cpsula de combusto. Esta cpsula colocada em uma bomba de oxignio contendo entre 25 e 30 atmosferas de oxignio. A bomba mergulhada em 2,000 g de gua em um calormetro adiabtico. Aps bomba e calormetro terem sido ajustados mesma temperatura, faz-se a ignio da amostra com um fusvel de metal. O aumento de temperatura medido sob condies adiabticas. Do equivalente hidrotrmico do calormetro multiplicado pelo aumento em temperatura, menos pequenas correes para a oxidao do fusvel e produo de cidos, calcula-se o valor calrico da amostra. EB = (TF - TI) x EHB - (CFQ x cal/ cm do fusvel) - ml de Na2CO3 onde: TF= Temperatura final TI= Temperatura inicial EHB= Equivalente hidrotrmico da bomba CFQ = Comprimento do fusvel queimado A energia bruta de alguns alimentos normalmente usados na alimentao animal esto descritos abaixo:
140
Fub de milho Farinha de trigo Soja Uria
4,43 kcal / grama 4,54 kcal / grama 5,52 kcal / grama 2,74 kcal / grama
Energia das fezes (EF): Uma amostra de fezes deve ser analisada seguindo o mesmo procedimento usado para avaliar amostras slidas e o clculo final baseado na seguinte frmula: EF = Peso seco das fezes x EB das fezes / unidade de peso seco.
Energia da urina (EU): Pode ser estimada pelo contedo em nitrognio da urina (NU) pela frmula: EU (kcal / kg) - 117 + 26 (NU%) Esta frmula foi desenvolvida com dados de bovinos e ovinos. Pode ser usado onde a quantidade de energia da urina representa uma pequena parte da energia total tal como na determinao da energia metabolizvel dos alimentos nas tabelas de composio dos alimentos. Caso seja necessrio uma determinao mais exata de energia da urina deve-se secar a amostra de urina e a energia determinada numa bomba calorimtrica de oxignio. A amostra de urina deve ser seca em contato com um combustvel slido de peso conhecido. Um pequeno becker de plstico serve tanto como recipiente de secagem como de combustvel slido. Uma amostra de 5 ml de urina pode ser pesada em becker de polietileno tarado. A secagem ao frio a preferida quando se dispuser desse tipo de secador. o prximo m mtodo preferido de secagem de amostras de urina em uma estufa vcuo 40C com mais de 700 mm de vcuo at que a amostra esteja quase seca. Quando no se puder secar dessa maneira, deve-se secar a urina em uma estufa comum 45C. A amostra no necessita estar completamente seca para ser queimada. A secagem em excesso resultar em perdas de energia desnecessria. Aps a secagem da amostra de urina no becker de plstico, a energia bruta determinada de acordo com o procedimento padro para outras amostras slidas. A energia bruta da urina calculada como se segue: EB da urina = (TF - TI) EHB - (CFQ x cal/cm de fusvel) - ml Na2CO3 - (PB x cal/g de poliestireno) (cal / g) Peso da amostra de urina em g onde: TF= Temperatura final TI= Temperatura inicial EHB= Equivalente hidrotrmico da bomba CFQ = Comprimento do fusvel queimado PB = Peso do becker de poliestireno
141
Coeficiente de digestibilidade aparente da EB: Este calculado atravs da seguinte frmula: 100 (Ebc - EF) / EBc.
1.3. CLCULOS BASEADOS NA COMPOSIO DO ALIMENTO: A. Energia bruta: possvel calcularmos os nveis de energia bruta de um alimento quando a sua composio conhecida. Na tabela 1 est descrita a composio de um determinado alimento A.
Tabela 1: Composio de um alimento A % 10,00 9,00 4,00 5,00 5,00 67,00 100,0
Umidade Protena Bruta Extrato Etreo Fibra bruta Resduo Mineral Extrativo no nitrogenado Total
Nas anlises de alimentos o extrato etreo representa os lipdeos totais e as fraes fibra e extrativo no nitrogenado representam glicdeos. O clculo da EB do alimento A est descrito na tabela 2.
Tabela 2: Nveis de energia bruta do alimento A Umidade Protena Bruta Extrato Etreo Fibra bruta Resduo Mineral Extrativo no nitrogenado Total % 10,00 9,00 X 4,00 X 5,00 X 5,00 67,00 X 100,0 Kcal/g 5,65 9,40 4,15 4,15 EB (kcal) 50,85 37,60 20,75 278,05 387,25
142
B. Energia digestvel: Desde que sejam conhecidos ou determinados os coeficientes de digestibilidade da matria seca total, ou dos princpios nutritivos tomados isoladamente, possvel calcular os nveis de ED. Considerando o mesmo alimento A (Tab. 1) e supondo os coeficientes de digestibilidade de 85% para protena bruta, 90% para extrato etreo, 5% para fibra bruta e 92% para extrativo no nitrogenado apresentamos na Tabela 3 os nveis de ED. Tabela 3: Nveis de energia digestvel do alimento A % Coeficiente de digestibilidad e % 85,00 90,00 5,00 92,00 Frao digerid a 7,65 3,60 0,25 61,64 Kcal/g ED (kcal)
Umidade Protena Bruta Extrato Etreo Fibra bruta Resduo Mineral Extrativo nitrogenado Total
10,00 9,00 4,00 5,00 5,00 no 67,00 100,0
x x x x
5,65 9,40 4,15 4,15
43,23 33,84 1,04 255,81 333,92
C. Energia metabolizvel : Se considerada a perda de 1,25 calorias por grama de protena absorvida vivel o clculo da energia metabolizvel de um alimento. Os nveis de energia metabolizvel do alimento A, j referido, esto apresentados na Tabela 4.
Tabela 4: Clculo de Energia metabolizvel para um alimento A % Coeficiente de digestibilidad e % 85,00 Frao digerid a 7,65 x Frao de E absorvida (Kcal/g) (5,65 -1,25) EM (kcal)
Umidade Protena Bruta
10,00 9,00
33,66
143
Extrato Etreo Fibra bruta Resduo Mineral Extrativo no nitrogenado Total
4,00 5,00 5,00 67,00 100,0
90,00 5,00 92,00
3,60 x 0,25 x 61,64 x
9,40 4,15 4,15
33,84 1,04 255,81 324,35
1.4. PREDIO DA ENERGIA PARTIR DE SEU CONTEDO FIBROSO: Os mtodos para estimar a energia devem ser rpidos, baratos e precisos, para serem adotados pelos laboratrios comerciais de testes de alimentos. Portanto, os laboratrios utilizam equaes empricas com base nos teores de fibra em detergente neutro (FDN) e fibra em detergente cido (FDA), para estimar o contedo de energia disponvel nos alimentos, principalmente em ruminantes. A relao vegetativa entre o contedo fibroso e a ED devida sua baixa digestibilidade, em relao frao nofibrosa (Van Soest, 1982). Diversos autores tm demonstrado que a digestibilidade de todos os componentes das forragens decrescem com o aumento do teor de FB na matria seca. Portanto, o contedo de fibra tem sido considerado o melhor preditor simples para todos os coeficientes de digestibilidade e a incluso de outros fatores aumenta relativamente pouco a acurcia das equaes de predio de digestiblidade. Vrios autores obtiveram equaes de predio para estimar a ED, EM e o contedo de outros nutrientes, para vrias espcies como eqinos, coelhos e sunos (Smolders et al., 1990; Maertens et al., 1987, Ortiz, 1989; Noblet & Perez, 1992). Segundo estes estudos encontrou-se uma relao negativa entre o contedo de fibra bruta (FB) e a digestibilidade aparente da matria orgnica, mas houve uma clara diferena entre a taxa de FB de forragens e de concentrados. De acordo com Almeida (1994), possvel predizer a ED dos alimentos usados para eqinos, partir da sua composio bromatolgica. As equaes que melhor predisseram os valores de ED para eqinos foram: ED = 5,0285 - 0,424 . FDA - 0,0144 . MO (r2 = 0,89) ED = 3,787 - 0,44 . FDA (r2 = 0,87)
1.5. VALOR AMIDO: No muito utilizado em nosso meio, apenas na Europa ainda utilizado. unidade energtica, podendo portanto, ser convertido em calor. O valor amido de um determinado alimento representa o nmero de unidades de amido puro que podem substituir 100 unidades desse alimento. 1.6. NIRS:
144
O desenvolvimento da tcnica de espectofotometria (NIRS) durante as ltimas duas dcadas tem gerado perspectivas para a utilizao de um mtodo de avaliao energtica barato, rpido e acurado em termos quantitativos e qualitativos. Alm disto uma tcnica que no necessita de reagentes qumicos e no produz poluentes. Este mtodo pode ser usado como mtodo de avaliao para classificao de amostras, facilitando o controle da composio qumica e dos valores energticos de alimentos usados na alimentao de bovinos, aves e sunos (Boever et al., 1994; Van der Meer et al., 1988; Valdes & Leeson, 1992). 2. MTODOS BIOLGICOS: Medidas calorimtricas e/ou anlises qumicas do alimento e das fezes so necessrias para determinar a digestibilidade. Levando-se em conta outras medidas energia da urina, energia dos gases produzidos (e perdidos), composio das descamaes epiteliais do trato gastrointestinal, etc... -, as estimativas vo ficando mais precisas e mais prximas do que realmente o alimento pode fornecer e, o animal, realmente utilizar e produzir. A digestibilidade pode ser avaliada in vitro ou in vivo. A avaliao da digestibilidade in vitro se baseia na produo de um ambiente que reproduza todas as condies encontradas in vivo, com a vantagem de ser um mtodo mais barato e prtico do que a digestibilidade in vivo. Se utiliza um tubo de ensaio e o pH do meio deve estar o mais prximo do pH do estmago da espcie estudada. Adiciona-se saliva artificial, a amostra e produz-se um ambiente anaerbio por meio de injeo de CO2 . Este material dever ser incubado em banho maria uma temperatura prxima do estmago da determinada espcie e aps este perodo o material dever ser centrifugado e o resduo dever ser separado. feita a mensurao da quantidade de matria seca do resduo e por diferena calculamos o quanto foi digerido. Atravs de equaes de regresso podemos obter correlaes entre valores de digestibilidade in vitro e in vivo. Os resultados in vitro so bem correlacionados com aqueles obtidos in vivo, mas geralmente os valores de digestibilidade in vitro so menores do que os in vivo. Os experimentos de digestibilidade in vivo so de alto custo e exigem que os animais permaneam um longo perodo consumindo grandes quantidades de alimentos, no sendo apropriados como metodologia rotineira, mas sim como testes de referncia. A digestibilidade pode ser considerada como real ou aparente. A digestibilidade real leva em conta as secrees endgenas (compostos nitrogenados, lipdeos e minerais), enquanto a aparente desconsidera essa frao. A determinao da digestibilidade in vivo pode ser feita da forma direta ou convencional, ou da forma indireta ou seja, atravs de indicadores. Os ensaios de digestibilidade so conduzidos com objetivo de avaliar diferentes dietas, quanto disponibilidade de nutrientes. Alguns pontos devem ser considerados quando se faz um experimento de digestibilidade:
� Adaptao do animal: O animal deve ser bem adaptado ao manejo e ao alimento
ser analisado. Esta adaptao feita de forma moderada consistindo de dois perodos: perodo de transio e de adaptao, que tem como objetivo alterar gradativamente a alimentao do campo para a alimentao do ensaio. Essa adaptao dura at que o animal atinja um nvel constante de consumo do alimento, aproximadamente 14 dias.
145
� Suprimento de gua: O consumo de gua interfere no consumo de alimento.
importante que a gua esteja disponvel para o animal, mas que no se misture a rao, para no alterar sua qualidade ou quantidade. Em muitos ensaios de digestibilidade os animais recebem gua duas ou trs vezes por dia. Esta freqncia est diretamente relacionada com a temperatura ambiente e a fase de vida do animal (lactao, vazia, gestante...). O efeito do consumo de gua na digestibilidade do alimento tem sido estudado mas para isto requer que o suprimento de gua seja controlado.
� Coleta de fezes: A durao deste perodo deve ser de 5-7 dias. Pode-se tambm
fazer perodos alternados de coletas. Antes do perodo da coleta devemos fazer um ensaio de consumo. Este ensaio ser baseado no seguinte clculo: Consumo = 100g de M.S./unidade de tamanho metablico (PV0,75). Durante o perodo de coleta devemos fazer a padronizao da ingesto. Essa padronizao da ingesto feita atravs do tamanho metablico, que igual ao peso do animal em kg, elevado 0,75. O perodo de coleta deve durar de cinco dez dias, para garantir a excreo fecal mdia constante, a fim de diminuir o efeito das variaes. As fezes devem ser coletadas diariamente, misturadas, e retiradas amostras representativas para cada animal, que sero secas temperatura entre 55 e 65C, para evitar a formao de substncias indesejveis (Van Soest, 1982).
� Separao da urina das fezes: A coleta de fezes tem que ser isenta de urina, para
no mascarar a digestibilidade. Por causa disso devemos usar machos de modo geral. No caso de aves, podemos fazer uma separao qumica com tratamentos especiais para separar o cido rico misturado nas fezes, ou ento fazer adaptaes cirrgicas. Os ensaios geralmente so feitos em gaiolas de metabolismo, que permitem a coleta de fezes e urina dos animais separadamente. Nas gaiolas normalmente existem funis para coletas de urina, sacolas ou caixas coletoras de fezes ou funis separadores.
� Uso de fmeas : No caso do uso de fmeas devero ser feita adaptaes para
desviar a urina (sondas, etc...)
� Seleo e preparao do alimento: Antes do incio do experimento uma quantidade
suficiente de cada alimento dever ser estocado. Os alimentos devero sempre ser oriundos da mesma fonte. Se no for possvel este alimento dever ser misturado em mesma proporo com o de outras fontes. O alimento a ser testado deve ser preparado e armazenado antes do incio do experimento, para evitar variaes na sua composio. Deve-se estabelecer o nvel de consumo durante um perodo preliminar, com durao mnima de duas semanas, para que os resduos da dieta anterior sejam eliminados do trato digestivo, bem como promover a adaptao dos animais dieta. A dieta oferecida de forma controlada para que no hajam sobras.
� Nmero de animais: Grindleu et al (1917) recomendou que no menos que trs
animais fossem usados em cada ensaio. Se for possvel devem ser usados cinco ou mais animais, de acordo com este autor. Resultados obtidos com quatro, cinco, seis ou mais animais so muito mais confiveis que aqueles obtidos com um ou dois, devido reduo da influncia da individualidade.
146
� Preparao dos animais: Aps a seleo dos animais, eles devem ser tratados para
parasitas internos e externos, vacinados contra algumas doenas contagiosas, identificados com brincos ou tatuagens e pesados individualmente. O manejo do animal dever ser tranqilo e freqente para que sua adaptao seja eficiente. Alguns fatores afetam a digestibilidade do alimento: Processamento do alimento (triturao, cozimento, peletizao, reconstituio de gros) Espcie animal: Respeitar as diferenas existentes entre monogstricos e ruminantes. Espcie vegetal Estdio fisiolgico da planta Idade do animal: Animais muito jovens e os muito velhos so piores. recomendado que se utilizem animais adultos jovens ou animais que esto em crescimento e atingindo a maturidade. Peso vivo: se reduz de 0,30-0,45 unidade a ED a cada 10 kg de incremento no peso vivo , em dietas contendo de 4-6% de fibra. Nvel de consumo: Quanto maior o consumo menor a digestibilidade, pois a digestibilidade depende da passagem do alimento pelo aparelho digestivo. Contedo de fibra: O teor de fibra aumenta a velocidade de passagem do alimento pelo intestino diminuindo a digestibilidade. Em monogstricos a digesto da fibra baixa o que influencia portanto na digestibilidade da dieta. A variao do coeficiente de digestibilidade da energia de um alimento utilizado para sunos est relacionado com a quantidade de polissacardeos no amilceos e de lignina. Em mdia o valor de coeficiente de digestibilidade da energia bruta varia de 70 90% (Tabela 5 e 6). Tabela 5: ED de alguns alimentos para sunos Alimento Fub de milho Gordura Glicdios solveis Energia digestvel (%) 100% 60-95% (varia com o grau de insaturao) 95-100%
A presena de polissacardeos no amilceos influenciam o coeficiente de utilizao intestinal daquele alimento e do valor do coeficiente de utilizao digestiva da dieta. Digesto parcial: um processo mais usado para ruminantes. Neste tipo de ensaio se estuda mais o local de absoro e no quantidade de absoro. O procedimento se baseia no abate do animal (durante o perodo de coleta) para coletar amostras nos diversos locais do intestino ou em adaptaes cirrgicas (fstulas) so feitas de maneira tal, que permitam coletar amostras em vrias partes do intestino. Geralmente o trato gastro intestinal do animal separado atravs de cnulas sendo a primeira fase chamada de pr-gstrica e depois a fase gstrica, e s vezes at o intestino grosso. Essas
147
separaes so feitas por cnulas reentrantes que exteriorizam o fluxo da digesta ou por fstulas simples (no fazerem amostragem quantitativa da digesta). Neste mtodo normalmente so utilizados indicadores. Os indicadores so substncias presentes no alimento ou que so adicionadas no alimento que so inertes, indigerveis, inabsorvveis e sem efeitos colaterais. Os indicadores podem ser classificados em internos (lignina, cromogenios, slios, nitrognio) que so aqueles existentes no prprio alimento, ou externos da fase slida (Cr2O3 , F2O3, Ru, Dy, Cs, etc...) e da fase lquida (polietileno glicol ). Esses indicadores externos possuem vrias formas. Exemplo de funcionamento dos indicadores: - Ingesto de 100 unidades de matria seca do alimento com 2% de indicador. - 60% foi digerido, logo 40% foi excretado. - Se esse indicador indigervel, as fezes apresentaro 5% de indicador: 100 unidades ingeridas com 2 unidades de indicador ----- 40 unidades excretadas com 2 unidades de indicador = 5%. - Para se calcular a indigestibilidade da amostra utilizada a seguinte frmula: IND = Conc. do indicador no alimento : Conc. do indicador nas fezes x 100
- Por diferena ser calculada a digestibilidade: Digestibilidade = 100 - Indigestibilidade Existem algumas consideraes que devem ser feitas em relao ao uso dos indicadores. - Indicadores externos (Cr2O3): Quando se trabalha com esses indicadores deve-se esperar algum tempo para que haja equilbrio entre a ingesto e excreo. Este equilbrio atingido de 6-8 dias. Porm, pode-se esperar mais para maior margem de segurana (10 dias). Aps atingido o equilbrio, inicia-se a fase de coleta das amostras. Os indicadores tem uma variao na excreo diria. Essa variao pode ser devido: - ao fato de o indicador no se misturar uniformemente com a digesta. - pode ser que a densidade do indicador seja maior que a do alimento em si. - pode tambm estar ligada ao hbito de ingesto do animal. - varia conforme o fornecimento do indicador (1 ou 2 vezes por dia). Essa variao pode ser diminuda fornecendo indicador de hora em hora. Tambm pode-se corrigir esta variao usando impugnao em papel ou associao de indicadores (indicadores da fase slida + indicadores da fase lquida).
148
Quando se mistura o indicador na rao pode-se diminuir a variao diria na excreo pelo animal. As desvantagens de se misturar o indicador que este pode no se misturar muito bem e pode ficar aderido s paredes do comedouro. Os cromognios vegetais so pigmentos de plantas tambm usados como indicadores internos. Geralmente so extrados com acetona 85%. Depois faz-se um padro de cromato de sdio da seguinte maneira: 10 unidades de cromognio = colorao de uma soluo com 5 mg de Na2CrO4 / 100 ml em um comprimento de onda de 406mM. Logo aps, compara-se a colorao do material extrado com o padro de cromato de sdio (processo calorimtrico). Esses cromognios vegetais no trazem problemas de equilbrio ingesto / excreo, variao diurna, etc... A desvantagem do uso dos cromognios vegetais que so parcialmente digeridos e absorvidos, por isso tem sido usados como ndices de digestibilidade O nitrognio outro componente do alimento utilizado como indicador, pois a digestibilidade da protena bruta relativamente constante. O nitrognio metablico fecal proporcional ingesto. A lignina que tambm pode ser utilizado como indicador dosada no alimento e nas fezes. Algumas desvantagens so: - parcialmente digerida, - necessria a determinao por anlise qumica. - Frmula qumica da lignina pouco conhecida e pouco estudada. V. CONVERSO DE UNIDADES: Atravs de frmulas podemos converter algumas unidades nos aproximando dos valores reais: 1. Converso de NDT em ED: Admitindo-se que 1 kg de NDT igual 4.409 kcal podemos calcular a ED de acordo com a frmula abaixo: ED (kcal / kg) = % NDT x 4,409 kcal 100 2. Converso de ED em EM: Para ruminantes: Para sunos: EM = 0,82 ED EM = ED x 96 - (0,202 x % PB) 100
VI. REQUERIMENTOS ENERGTICOS DOS ANIMAIS: Energia para metabolismo basal:
149
Grande parte dos valores que expressam necessidade, so deduzidos do peso metablico, ou seja, do peso especfico (peso vivo) elevado potncia 0,75. Em virtude de que a superfcie do corpo do animal ser uma funo do peso, pode-se estabelecer o clculo da necessidade energtica partir da seguinte relao: Cal = a Pvb; onde: Cal = Calorificao por 24 horas PV = Peso vivo a e b = Coeficientes para os quais Brody encontrou os valores de 70,5 para a e 0,73 para b, vlidos para animais com pesos entre 20 g e 600 kg. Posteriormente Kleiber estimou que em lugar de 0,73 para o coeficiente b, o valor mais adequado era de 0,75. A equao para definir o gasto energtico mnimo (metabolismo basal) ficou corrigida para: Cal = 70,5 x PV 0,75 e define o gasto energtico mnimo dos processos autnomos do organismo, representados pelas reaes endo e exotrmicas, ou seja, de um animal em estado de ps-absoro (jejum), em repouso e em ambiente termicamente neutro. Energia de mantena: Seguindo-se ao metabolismo basal a necessidade energtica imediatamente superior corresponde ao estado de manuteno ou mantena, e que se refere a um indivduo adulto da qual no se exige um gasto excessivo em termorregulao, nem um gasto ligado alimentao anormal, bem como sem apresentar produo. Deve estar em estado hgido adequado, desenvolvendo a atividade normal nas condies acima descritas. equao acima deve-se acrescentar um coeficiente para converter a necessidade calrica em repouso para aquela de manuteno: Cal = a x 70,5 x PV0,75 onde a apresenta os valores medidos de 2 para os herbvoros e de 1,4 para os onvoros. Para aves esta equao citada acima no aplicvel, uma vez que estas apresentam uma temperatura corporal mais alta do que a dos mamferos, exigindo maior gasto de energia para sua manuteno. Para aves adultas a equao aplicvel para o clculo do gasto energtico basal a seguinte: EL = 83 x PV0,75 A necessidade energtica de manuteno , pois, o ponto de partida para aquelas de produo, crescimento (produo de carne), produo de leite, ovos, l, etc...
150
Energia de crescimento: O crescimento representa uma variao de peso contnuo, no sentido do aumento. Portanto, no se pode considerar um animal em crescimento como, em um dado momento, estando em manuteno, pois a cobertura de suas necessidades energticas deve atender a manuteno mais o aumento de peso pretendido na continuidade, que constante, observando uma curva sigmidea prpria de cada espcie. Igualmente, o crescimento, dos diferentes tecidos no ocorre com a mesma rapidez, podendo-se considerar que o crescimento dos msculos e do esqueleto representam cerca de 48 vezes para os primeiros e apenas cerca da metade desta proporo para o ltimo, desde o nascimento at a maturidade. Ao se observar uma curva de crescimento, nota-se a existncia de dois segmentos, o primeiro, crescente, vai do nascimento at a puberdade enquanto que o segundo, decrescente, vai da puberdade ao estado adulto. As curvas de crescimento prprias de cada espcie, ou mesmo de cada linhagem, so tomadas como padres para compar-las com o atendimento das necessidades nutricionais. Estes padres so obtidos em testes biolgicos, onde se verifica a ingesto de energia, correlacionada com os outros nutrientes, necessria para atingir o peso mximo dentro do estabelecido para a linhagem. Pode-se orientar o teste para o conhecimento, tambm, da energia necessria para a produo de um quilograma de peso, a fim de, comparativamente com a curva de crescimento padro, estabelecer a eficincia alimentar mais econmica em face dos preos de mercado. Energia para produo de leite: Para a produo de leite, conhecendo-se a composio mdia do mesmo, fcil deduzir-se o gasto energtico para a produo de um quilograma do produto. Multiplicando os constituintes do leite pelos seus correspondentes calricos brutos (ex: gordura x 9,4 etc...), teremos a energia perdida atravs do mesmo. Uma vez que o teor em gordura a maior varivel dos princpios nutritivos, pode-se padronizar o clculo para leite com 4% de gordura, aplicando-se a frmula de Gaines: L = 0,4 + 0,15 x , onde L a quantidade de leite corrigido para 4% de EE, comparativamente 1 kg deste com x% de gordura. Evidentemente, alm da energia perdida para cada kg de leite (energia lquida leite), existe um gasto energtico para produo de 1 kg do produto. Forbes e Kriss calcularam que o gasto energtico para a produo de leite, em energia metabolizvel 1,67 vezes o gasto energtico prprio do leite produzido. VII. BALANO ENERGTICO A mensurao do balano nutricional tem como finalidades verificar a eficincia de utilizao do alimento, determinao das exigncias nutricionais, mensurao do valor energtico dos alimentos e do valor biolgico das protenas. Existem duas tcnicas de avaliao do balano: - Balano da matria: Balano dos nutrientes que podem ser pesados, coletados, etc... - Balano da energia: Envolve as tradicioais perdas insensveis do corpo (calor, gases, etc...). Para a avaliao do balano necessrio a utilizao de equipamentos sofisticados. No balano energtico mede-se a ingesto de energia bruta, a excreo de energia bruta nas fezes, urina e leite, produo de calor dos animais e produo de
151
gases. Dois processos so usado na mensurao do calor dos animais: mtodos diretos ou indiretos. - Indiretos: Abate comparativo. Se formam dois lotes de animais (bem uniformes), sendo que um lote abatido e o outro submetido a um perodo de alimentao. No lote abatido, dosamos o teor de energia do corpo dos animais. - Diretos (tcnicas calorimtricas): Pode ser feita a calorimetria direta ou indireta. Para a calorimetria direta podemos usar os calormetros de gelo, adiabticos, ou de gradiente. Na calorimetria indireta usam-se cmaras de respirao, mscaras faciais, ou cnulas traqueais. Nestes casos as trocas gasosas so mensuradas. Existem cmaras de respirao de circuito aberto, com renovao de ar e de circuito fechado, onde o mesmo ar, aps a retirada de CO2, metano e outros gases reaproveitado. Nessa cmara mede-se o CO2 produzido, O2 consumido, metano produzido, etc.. e a se calcula os chamados quocientes respiratrios (QR). QR = Vol. de CO2 produzido : Vol. de O2 consumido. Atravs do QR, pode-se estimar a produo de calor dos animais. REFERNCIAS BIBLIOGRFICAS ALMEIDA, M.I.V. Predio da energia digestvel de dietas para eqinos partir de seu contedo fibroso. 1994, Escola de Veterinria da UFMG, Belo Horizonte, 110p., Tese. ANDRIGUETTO, J.M.; PERLY, L.; MINARDI, I.; GEMAEL, A. et al. Avaliao do valor energtico dos alimentos. In: Nutrio Animal, 257 - 267 (I), So Paulo, Nobel, 1982, 389p., 1982. BERGMAN, E.N. Energy contribuitions of volatile fatty acids from the gastrointestinal trait in various species. Physiological Reviews, 70 (2), 1990. BOEVER J.L., COTTYN, B.E., VANACKER, J.M., BOULQU, Ch. V. The use of NIRS to predict the chemical composition and the energy value of compound feeds for cattle. Animal Feed Science and Technology, 51, 243-253, 1995. DALE, N. The metabolizable energy of wheat by products. J. Appl. Poultry Res., 5, 105- 108, 1996 HARRIS, L.E. In: Compilao de dados analticos e biolgicos para o preparo de tabelas de composio de alimentos para uso nos trpicos da amrica latina. I. Procedimentos para descrever e analisar amostras de alimentos e registro dos dados na fonte de informaes. 1970, University of Florida, Gainesville, USA, 1970. HARRY, W. T. In: The Scientific Feeding of Chickens, 1955, The Interstate, Illinois, 297p., 1955.
152
MAERTENS, L., MOERMANS, R., DeGROOTE, G. Estimation de la teneur en nergie des aliments pous lapins. Revue de lAgriculture,v.40,n.5, p.1205-1216, 1987. NOBLET, J.; PEREZ, J.M. Prediction of digestibility of nutrients and energy values of pig diets from chemical analysis. J. Anim. Sci., v.71, p.3389 3398, 1992. ORTIZ, V. Determinacion de las necessidades energeticas y valoracion de piensos para conejos en crecimento por el metodo de calorimetria indirecta. Madrid: Universidad Politecnica de Madrid, 1989, 100 p. (Tese de mestrado em Nutrio Animal). ROSTAGNO, H.S., SILVA, D.J., COSTA, P.M.A., FONSECA, J.B. et al. In: Composio de alimentos e exigncias nutricionais de aves e sunos (tabelas brasileiras), 1983, Viosa, UFV, Impr. Univ., 60 p., 1983. SCHNEIDER, B.H. & FIATT, W.P. In: The evaluation of feeds through digestibility experiments. 1975, University of Georgia Press, 423p., 1975. SILVA, D.J. Determinao da energia bruta - Uso da bomba calorimtrica. In: SILVA, D.J. Anlise de alimentos (mtodos qumicos e biolgicos), p. 25-32, Viosa, U.F.V., Impr. Univ., 1981. SMOLDERS, E.A.A.A., STEG, A., HINDLE, V.A. Organic matter digestibility in horses and its prediction. Netherlands J. Agric. Sci., v.38, p.435-447, 1990. VALDES, E.V.; LEESON, S. Near infrared reflectance analysis as a method to measure metabolizable energy in complete poultry feeds. Poult.Sci., v.71, p. 1179 - 1187, 1992. VAN DER MEER, J. M., VEDDER, H., WEVER,G. NIRS for the prediction of organic matter and energy digestibility in pig feeds. In Vitro Newslett., v.4, p.25-26, 1988. VAN SOEST, P.J. Nutritional Ecology of the Ruminant. Corvalllis, O & B. Books, 1982. 373p.
VITAMINAS INTRODUO De modo geral, chamase vitamina uma substncia orgnica que preenche os seguintes quesitos: 1 Ser um componente de um alimento natural, mas no sendo carboidrato, lipdio ou protena; 2 Estar presente em concentraes muito pequenas na maioria dos alimentos;
153
3 Essencial para o metabolismo normal dos animais e conseqentemente necessria para a sade e funes fisiolgicas como crescimento, desenvolvimento, manuteno e reproduo; 4 Causar uma doena ou sndrome especfica, quando ausente na dieta ou quando mal absorvida ou utilizada; 5 Quando biossintetizvel, no pode o ser em quantidades suficientes para preencher as necessidades fisiolgicas, devendo conseqentemente ser obtida a partir da dieta. A maioria das vitaminas tradicionais, tais como A, E, K, B1, B6 e B12, riboflavina, cido flico, cido pantotnico e biotina satisfazem cada um desses requisitos. Por sua vez, as vitaminas C e D, a niacina e a colina so consideradas vitaminas somente em contextos bem definidos. Por exemplo, a vitamina C (cido ascrbico) metabolicamente sintetizada pela maioria das espcies a partir da glicose na via do cido glicurnico. Somente poucas espcies como os seres humanos, as cobaias, alguns peixes, etc. por falta da enzima L-gulonolactona oxidase no sintetizam o cido ascrbico, requerendo, portanto esta vitamina na sua dieta. A vitamina D por sua vez realmente vitamina em condies de ausncia de luz solar (sistemas de confinamento modernos); pois as radiaes ultravioletas da luz solar permitem a biossntese de vitamina D a partir dos precursores. A niacina metabolicamente sintetizada a partir do aminocido essencial triptofano. Contudo, a eficincia desta converso to baixa que a maioria das espcies (especialmente o gato, os peixes, pato e marreco) necessitam de niacina diettica quando os teores de triptfano recebidos so simplesmente aqueles das misturas dietticas comuns. A colina, que metabolicamente gerada pela aminao e subsequente metilao da serina no biossintetizada em quantidades suficientes para satisfazer as exigncias do rpido crescimento dos pintos e de outras aves domsticas jovens. Neste caso, a colina uma vitamina. VITAMINA A 1 As formas da vitamina A A expresso "atividade vitamnica A" uma caracterizao genrica dos derivados da -ionona com atividade biolgica do tudo-trans-retinol. Nas plantas, esta atividade est presente somente na forma dos carotenides precursores do referido transretinol. O mais ativo desses carotenides o -caroteno. Os alimentos (matrias primas) de origem animal contm carotenides e/ou retinides. Os mais significantes retinides do metabolismo animal so o lcool ( tudo-trans-retinol ), os aldedos (11-cis retinal e 11-cis-3 dehidroretinal), o cido retinico (tudo-trans), bem como os retinil esteres (principalmente o retinil palmitato) e retinil--glicurondio. 2 Absoro e Metabolismo Vrias formas de vitamina A e carotenides so absorvidos principalmente junto com lipdios. Os carotenides so normalmente convertidos em retinol na mucosa intestinal; mas provavelmente tambm no fgado e outros rgos, especialmente nas especies que possuem gordura amarelada tais como os bovinos e aves domsticas. Os retinil esteres da dieta so hidrolisados a retinol no intestino; e absorvidos na forma de lcool (retinol) livre que em seguida reesterificado na prpria mucosa. Nos mamferos, os retinil esteres so transportados principalmente ligados aos qulilomicrons da linfa,
154
para o fgado onde so hidrolisados a retinol e reesterificados antes de serem estocados. Quando da mobilizao da vitamina A do fgado, o ester hidrolisado liberando o retinol. O retinol sai do fgado ligado a uma protena, a qual o transporta para os tecidos. Aps a utilizao do retinol pelos vrios tecidos, o mesmo pode ser oxidado para aldedo (retinal) o qual pode ser mais tarde oxidado irreversivelmente a cido retinico. Na hipervitaminose A, a toxidez se deve principalmente ao cido retinico, bem como ao retinal em face de sua fcil converso no organismo a cido retinico. Entretanto, o cido retinico um alternativo metablico com funo de vitamina A, exceto para as funes de viso e reproduo. A principal via de excreo da vitamina A biliar, atravs da conjugao do cido retinico com o cido glicurnico, sendo que o glicurondio formado eliminado na bile. A circulao entero-heptica provavelmente serve para poupar a vitamina A ao invs da excreo fecal. Pequenas quantidades de glicurondio e metablitos finais no identificados so excretados via urina. 3 Funes A vitamina A possui um papel bioqumico na viso noturna. No olho, o trans-retinol transformado em 4-cis-retinol, o qual oxidado a 4-cis-retinal. O 4-cis-retinal combinase com a protena opsina para formar a rodopsina tambm conhecida como prpura visual. Quando a rodopsina exposta luz, o 4-cis-retinal, que est ligado com a opsina, convertido em trans-retinal, transformando-se assim a rodopsina em opsina livre e trans-retinal. A energia liberada nesta reao transmitida ao crebro atravs do nervo ptico. Finalmente, o trans-retinal reduzido reversivelmente a trans-retinol, ou oxidado irreversivelmente a cido retinico. Outra literatura (Vitamin Tolerance of Animals) relata uma verso ligeiramente diferente, afirmando que no olho o retinol oxidado a retinal, com a subseqente isomerizao do trans-retinal a 11cis-retinal, o qual se combina com a opsina para formar a rodopsina. Os mecanismos moleculares de atuao da vitamina A no crescimento, reproduo e integridade epitelial ainda permanecem parcialmente escuros. Mas a hiptese mais amplamente aceita so o papel na sntese de glicoprotenas que controlariam a diferenciao celular, e o envolvimento no controle da expresso gnica. Alm da proteo do epitlio germinativo nos machos e epitlio uterino nas fmeas, a vitamina A promove um bom desenvolvimento embrionrio. A vitamina A atua tambm no desenvolvimento do sistema nervoso. O -caroteno possui algumas funes no relacionadas com sua atividade provitamnica A. O corpo lteo das vacas leiteiras contm uma considervel concentrao de -caroteno. Baixos nveis do mesmo no plasma de bovinos (vacas) esto relacionados com problemas reprodutivos. VITAMINA D 1 As formas da vitamina D As diferentes formas de vitamina D (esteris) utilizadas na medicina humana e animal correspondem variadas potncias biolgicas relativas.
155
A vitamina D2 menos potente nas aves do que nos mamferos, ao passo que a D3 potente em ambas as espcies.
2 Absoro, metabolismo e funes Por ser lipossolvel, a vitamina D absorvida junto com outras gorduras via quilomicrons no sistema linftico dos mamferos ou na circulao portal das aves e dos peixes. As duas principais formas de vitamina D so o colecalciferol (Vit. D3) e o ergocalciferol (Vit. D2). O colecalciferol sintetizado na pele dos animais a partir do colesterol. Nesse processo o colesterol oxidado a 7-dehidrocolesterol, que em presena da luz ultravioleta se transforma em colecalciferol. O ergocalciferol por sua vez sintetizado exclusivamente pelos vegetais, a partir do ergosterol, tambm em presena das radiaes ultravioletas. A vitamina D absorvida ou biosintetizada transportada atravs da circulao para o fgado, rim e mucosa intestinal onde ocorre a primeira hidroxilao gerando a 25hidroxivitamina D. A segunda hidroxilao acontece no rim, resultando na forma metabolicamente ativa da vitamina D (1, 25-dihidroxicolecalciferol para vit. D3, ou 1, 25-dihidroxiergocalciferol no caso da vit. D2). A 1, 25-dihidroxivit D, o paratormnio (PHT) e a calcitonina regulam a calcemia. Uma queda na calcemia causa aumento da secreo de PTH, o qual estimula a biosntese de 1, 25-dihidroxivit D, que determina um aumento da absoro intestinal de clcio e da reabsoro do clcio sseo. De fato, a vit. D promove a sntese da protena transportadora de clcio, a qual realiza o transporte ativo deste mineral. Por outro lado, o aumento da calcemia resulta na diminuio da secreo de PTH e aumento da secreo de calcitonina. Isso resulta na reduo da produo de 1, 25-dihidroxivit D, causando reduo da absoro intestinal de clcio e da reabsoro ssea. Outros fatores tambm podem influenciar a sntese de 1, 25-dihidroxivit D. VITAMINA E 1 As formas da vitamina E O nome vitamina E a designao genrica dos derivados do 6-cromanol com atividade biolgica do -tocoferol. Na tabela a seguir est a lista das referidas substncias com suas potncias relativas e fontes. 2 Absoro e metabolismo A vitamina E absorvida na forma de lcool livre (tocoferol). Ela insolvel no meio aquoso da luz intestinal, sendo solvel somente na gordura. A sua absoro bem como a de outros nutrientes exclusivamente ou predominantemente liposolveis depende, portanto, da sua solubilizao micelar, isto , depende da formao de micelas. Consequentemente, o no funcionamento do pncreas bem como uma hipoproduo biliar prejudica essa absoro. Nos mamferos, o tocoferol absorvido transportado pelos quilomicrons da circulao linftica para o fgado, e subsequentemente para a circulao geral por lipoprotenas de densidade muito baixa (VLDL). Nas aves e nos peixes, os lipdios
156
absorvidos seguem para o fgado pela circulao portal. O fgado e provavelmente todos os tecidos extrahepticos recebem a vitamina E das VLDL. A vitamina E est presente nos tecidos na forma de lcool livre. O tocoferol age cedendo hidrognio para a reduo dos radicais livres. Ele cede o hidrognio do seu grupo 6-OH, se transformando em 8--hidroxitocoferona ou 8-alkoxi--tocoferona. Pela subsequente hidrlise, as tocoferonas se transformam irreversivelmente em tocoferil quinonas. 3 Funes A vitamina E um antioxidante biolgico, funcionando tambm in vitro como timo antioxidante. O selnio ( por ser um componente fundamental da enzima glutationa peroxidase) e a vitamina E fazem parte do sistema defensivo antioxidante, da a interrelao entre vit. E e selnio. Este sistema protege as clulas contra os danos causados pelo oxignio reativo e outros radicais livres que oxidam os fosfolipdios insaturados das membranas e/ou as protenas crticas. Aqui, o mecanismo real de ao da vitamina E o fornecimento de hidrognio na reao que leva reduo do oxignio reativo e de outros radicais livres. Acredita-se que a vitamina E por ser lipossolvel protege as membranas celulares (pois estas so fundamentalmente lipdicas); embora atue tambm no cistosol e na matriz mitocondrial. Ao contrrio, a glutationa peroxidase protege somente os componentes citoslicos e da matriz mitocondrial. por isso que dentre os sndromes de natureza carencial referentes ao sistema antioxidante, alguns respondem somente vitamina E ou a antioxidantes sintticos liposolveis enquanto outros podem ser resolvidos tanto com a vit. E quanto com o selnio. Supe-se que os primeiros so causados por leses nas membranas e os ltimos mencionados devidos a leses nas membranas e nos componentes celulares solveis no citosol ou na matriz mitocondrial. A vitamina E assegura a respirao normal dos tecidos, possibilitando por um mecanismo pouco elucidado o funcionamento da redutase do citocromo C, e a proteo antioxidativa das estruturas lipdicas mitocondriais. Ela necessria para as reaes de fosforilao, especialmente naquelas que geram molculas de alta energia. Participa no metabolismo dos cidos nuclicos, na sntese da vitamina C, na sntese da ubiquinona e no metabolismo dos aminocidos sulfurados. importante na funo reprodutiva. Essencial para a manuteno da funo testicular protegendo do epitlio germinativo. Sua carncia nas fmeas causa parada do crescimento do feto, sua reteno e reabsoro. Nas aves, a deficincia prolongada leva esterilidade nos machos; e morte embrionria. A vitamina E protege o mesoderma e os tecidos dele originados. Atua no metabolismo da cistena. VITAMINA K 1 As formas da vitamina K Os componentes com atividade vitamnica K possuem o anel 2-metil-1,4naftoquinona. O termo vitamina K genrico, designando o 2-metil-1,4-naftoquinona e todos os derivados gerados por modificao na posio (carbono)3, que exibem uma
157
atividade antihemorrgica em animais deficientes em vitamina K. A vitamina K1 ou filoquinona sintetizada pelas plantas. A vitamina K2 ou menaquinona designa uma srie de derivados sintetizados por bactrias. Por sua vez, a vit. K3 tambm conhecida por menadiona uma forma sinttica. o prprio anel 2-metil-1,4 naftoquinona. Para se tornar ativa a vitamina K3 alkilada por enzimas presentes no tecido dos mamferos. 2 Absoro A vitamina K absorvida do intestino seguindo a via linftica nos mamferos. A absoro requer a presena de gorduras, bem como das secrees biliar e pancretica. 3 Funo Na deficincia de vitamina K ou na presena de seus antagonistas, os animais apresentam hemorragias espontneas.Isso se deve falta de protrombina normal. No organismo a protrombina recm formada se encontra numa forma anormal. Ela transformada em protrombina normal mediante um processo de carboxilao catalizada por uma carboxilase cujo cofator a vitamina K. A protrombina normal possui os primeiros 10 glutamatos da regio aminoterminal carboxilados. Isto faz com que no mecanismo de coagulao sangunea, a ligao da protrombina com o clcio seja forte, o que promove a ancora desta protena (protrombina) s membranas fosfolipdicas das plaquetas, resultando numa ntima proximidade da protrombina com os fatores X e V que participam da sua ativao. Na seqncia ocorre separao do fragmento aminoterminal (ativao). A trombina resultante liberada no plasma promove a ativao do fibrinognio (transformao em monmero de fibrina). Na falta de vitamina K ou na presena de seus antagonistas, ocorre sntese da protrombina, entretanto, esta permanece na forma anormal. Ela praticamente no se liga ao clcio e consequentemente ineficaz no processo de coagulao sangunea. Radicais especficos de glutamato nos fatores VII, IX e X tambm so carboxilados, formando centros com alta afinidade por Ca+. Esta carboxilao depende da vitamina K. No que diz respeito ao fator IX, a ligao do clcio promove a sua ativao. A protrombina anormal assim como os fatores VII, IX e X no carboxilados so denominados por certas literaturas de protenas precursoras dos fatores completos ( formas carboxiladas ) � DUKES (vit. K). VITAMINA B1 1- As formas da vitamina A vitamina B1 ou tiamina encontrada na maioria dos tecidos animais predominantemente nas formas fosforiladas (como exemplos temos as tiaminas mono, di e trifosfato). Nos cereais e nas leguminosas se encontra numa forma no fosforilada. O hidrocloreto e o mononitrato de tiamina so sintetizados para uso nas raes animais. Conseqentemente, a tiamina encontrada nos alimentos nas seguintes formas: tiamina livre, tiamina fosforilada e tiamina em complexos fosfo-proticos.
158
2 Absoro e metabolismo No tubo gastrintestinal, as formas de tiamina ligada a outros compostos so clivadas e a tiamina livre absorvida principalmente na poro proximal do intestino delgado. A distribuio tecidual da tiamina parece ser bastante uniforme, sendo encontradas as mais altas concentraes no fgado e no rim. No organismo humano, cerca de 80% da tiamina existe na forma de tiamina pirofosfato (TPP), 10% como trifosfato e o restante consiste do monofosfato. A timina uma das vitaminas de menor concentrao (estoque) no organismo. Os mamferos podem esgotar suas reservas corporais de vitamina B1 dentro de uma a duas semanas. Entretanto, o suno uma exceo a esta regra geral, pois tem a possibilidade de estocar grandes quantidades de tiamina no msculo esqueltico. Nos mamferos, o excesso de tiamina eliminado principalmente atravs da urina sem sofrer alterao. Entretanto, um certo nmero de diferentes metablitos da tiamina foram encontrados na urina humana e de rato.
3 - Funes da tiamina. A forma metabolicamente ativa da tiamina a tiamina pirofosfato (TPP), que uma coenzima. A TPP cofator na descarboxilao oxidativa de -cetocidos como o cido pirvico e o cido -cetoglutrico. A transformao do cido pirvico em acetil CoA pela descarboxilao oxidativa, gera um grupamento acetil, que pode servir para sintetizar molculas como a acetilcolina, que um neurotransmissor; razo pela qual a deficincia de tiamina pode causar distrbios neuromusculares. A tiamina pirofosfato atua tambm como coenzima da transcetolase no ciclo das pentoses-fosfato. RIBOFLAVINA (VITAMINA B2) 1 As formas da vitamina As formas desta vitamina nos alimentos seriam a riboflavina ou as coenzimas FMN (flavina mononucleotdio) e FAD (flavina adenina dinucleotdio), que so as formas predominantes nos tecidos de mamferos. A forma suplementar para as dietas geralmente a riboflavina, embora alguns pesquisadores tenham experimentado o uso de riboflavinato de sdio. Esta forma mais hidrossolvel do que a riboflavina.
2 Absoro e metabolismo A riboflavina sintetizada pela flora intestinal, principalmente nos ruminantes onde h tambm sntese a nvel ruminal. O tempo de trnsito do bolo influencia a absoro. Contudo, no se sabe a eficincia da absoro da frao sintetizada pela microflora nos monogstricos. A riboflavina na circulao est ligada a protenas, inclusive imunoglobulinas.
159
A absoro desta vitamina ocorre no intestino delgado. As reservas parecem ser pequenas. A vitamina B2 rapidamente excretada na urina, o que explica o fato dos animais necessitarem constantemente da mesma. 3 Funes A riboflavina atua na transferncia intermediria de eltrons nas reaes metablicas de oxi-reduo sob forma de duas coenzimas, FMN e FAD. Estas coenzimas funcionam com um grande nmero de oxidases e desidrogenases importantes no metabolismo normal. As enzimas que usam o FMN incluem a glicose oxidase, L-aminocido oxidase e lactodesidrogenase. As que usam o FAD incluem a Daminocido oxidase, a desidrogenase succnica, as desidrogenases de acil CoA, a Lgulonalactona desidrogenase, a -glicerofosfato desidrogenase e a glutationa redutase. Esta vitamina possui tambm um papel importante na sntese de polissacardios. Nos estados de estresse quando a necessidade de hormnios adrenais aumenta, a riboflavina importante, pois parece necessria para a sntese dos referidos hormnios. VITAMINA B12 Isolada como fator antianemia perniciosa, a vitamina B12 apresenta-se na forma de cristais avermelhados higroscpicos, solveis em gua, lcool, cido actico, insolveis no clorofrmio e no ter. A cianocobalamina (vit. B12) possui um on CN cuja substituio por OH gera a hidroxicobalamina, que outra forma de vitamina B12. A sua presena nos tecidos animais devida ingesto de produtos de origem animal ou sntese microbiana a nvel do aparelho digestivo. Vrios derivados da cianocobalamina, incluindo a hidroxicobalamina e a nitritocobalamina tambm possuem atividade de vitamina B12. 1- Absoro e funes A sua absoro no aparelho digestivo depende da presena do fator intrnseco, uma mucoprotena, a qual encontrada no suco gstrico. A absoro ocorre unicamente no leo, conforme a dosagem. Esta vitamina transportada no sangue combinada a duas protenas (transcobalaminas I e II) e estocada no fgado. Atua juntamente com o cido flico na biossntese de grupamentos metil lbeis. Esses grupamentos so necessrios para a biossntese das bases pricas e pirimidnicas, da metionina a partir da homocistena, e da colina. A vitamina B12 necessria para o metabolismo glicdico como coenzima da enzima desidrogenase do gliceraldedo-3-fosfato. Atua ainda no metabolismo lipdico via seus efeitos nos tiis (mantm os grupamentos sulfidrila na forma reduzida). As coenzimas cobamida muitas vezes necessitam interagir com outros nutrientes, tais como a riboflavina, o cido nicotnico e o magnsio. Essas coenzimas esto relacionadas com a biossntese de aminocidos, bem como com o metabolismo do cido ascrbico e com a funo da tireide.
160
COLINA A colina atua na regulao de processos metablicos como um dos principais fornecedores de grupamentos metil lbeis para os processos de transmetilao. Funciona tambm como elemento plstico na constituio de componentes citoplasmticos e humorais. A colina possui funes bsica e de lcool, podendo a funo lcool ser acetilada formando a acetil-colina (neurotransmissor). A colina encontrada em quase todos os alimentos s vezes em estado livre, mas na maioria das vezes combinada sob a forma de lecitina. Quando os grupos metil lbeis esto presentes no organismo em quantidades adequadas, a colina pode ser sintetizada em nvel suficiente para atender s necessidades do organismo. Outras funes Participa da sntese fosfolipdica. Ao lipotrpica, prevenindo o acmulo anormal de gordura no fgado.
VITAMINA C Diversas substncias tm atividade de vitamina C, sendo que a mais importante o cido ascrbico. Essa vitamina atua como importante substncia redox do metabolismo celular. O cido 1-ascrbico e seu produto de oxidao, o cido dehidro-1-ascrbico formam o sistema redox. A maioria das espcies animais capaz de sintetizar a vitamina C no seu organismo, com exceo do macaco, do homem e da cobaia. A sua absoro em condies normais fcil e rpida. O seu produto final de catabolismo o cido oxlico, que excretado pela urina. Funes Algumas reaes nas quais a vitamina C participa so:
� � �
Sntese de corticosterides, principalmente nas condies de estresse. Evita a oxidao metablica de certos aminocidos inclusive a tirosina. Atua na transferncia do ferro da protena plasmtica (transferrina) para a protena celular (ferritina) favorecendo a estocagem do ferro no organismo. Ainda, a vitamina C importante para na absoro do ferro no aparelho digestivo. Atua como fator necessrio para a ativao do cido flico no fgado, o transformando em cido folnico (forma metabolicamente ativa).
161
� � �
� � �
Efeito estimulante na atividade fagocitria dos luccitos, nas funes do sistema retculo-endotelial e na formao de anticorpos. Atua no processo de hidroxilao da prolina em hidroxiprolina, componente essencial das fibras colgenas; O papel principal mais especfico do cido ascrbico parece ser a sua participao na formao da matriz protica dos ossos e dos dentes, bem como na sntese de colgeno. Necessria para a formao das substncias intercelulares, interfibrilares, compostos de mucopolissacardeos. Parece reduzir os nveis de colesterol sanguneo. Atua na recuperao (reciclagem) da vitamina E aps que esta reduz uma radical livre.
NIACINA 1- As formas da vitamina Niacina o termo genrico para designar os cidos piridino-3 carboxlicos e seus derivados, que possuem a atividade biolgica da amida do cido nicotnico, ou seja, da nicotinamida. Dentre os compostos do grupo da niacina, o cido nicotnico e a nicotinamida so os biologicamente mais potentes. A niacina existe nos alimentos em geral, quer de origem vegetal ou animal. Os cereais constituem a mais importante fonte de niacina na maioria das dietas dos animais. 2- Absoro e metabolismo O cido nicotnico e a nicotinamida so absorvidos quase completamente por difuso simples atravs da mucosa intestinal. Certos pesquisadores acreditam que o cido nicotnico uma vez absorvido transformado na mucosa intestinal em nicotinamida, a qual captada pelos tecidos e transformada nas coenzimas NAD e NADP. Outros sustentam que o cido nicotnico tambm pode ser diretamente utilizado na biossntese de NAD e NADP, sendo que neste caso a transformao do cido em nicotinamida ocorre somente na ltima etapa, num composto intermedirio de biossntese. Outros intermedirios de biossntese de NAD e NADP so a nicotinamida mononucleotdio (NMN) quando a sntese se faz a partir da amida, ou o cido nicotnico mononucleotdio quando o precursor o cido nicotnico. A niacina pode ser biossintetizada a partir do triptofano, sendo que a eficincia dessa converso varia conforme a espcie animal. O gato praticamente no sintetiza. Os outros fatores que afetam a converso incluem o nvel de leucina, da protena total e a piridoxina. A niacina e seus metablitos so excretados na urina, principalmente em caso de ingesto de altas doses. 3-Funes A base bioqumica dos diversos efeitos da deficincia de niacina envolve uma srie de reaes metablicas das quais participa a nicotinamida. Estas incluem cerca de 35
162
reaes de oxidoreduo das quais participa a NADH (nicotinamida adenina dinucleotdio) ou a NADPH (nicotinamida adenina dinucleotdio fosfato) atuando como transportadores de dois eltrons. NADH transporta eltrons dos intermedirios metbolicos para a cadeia respiratria, ao passo que NADH e NADPH servem como redutores num grande nmero de biossnteses. Consequentemente, a nicotinamida tem funes fisiolgicas crticas na respirao mitocondrial e no metabolismo dos carboidratos, lpidios e aminocidos. CIDO PANTOTNICO 1 - As formas da vitamina O cido pantotnico o cido pantico ligado -alanina por uma funo amida. Boa parte do cido pantotnico nos tecidos est representada pelas suas formas coenzimticas. Todas essas coenzimas possuem a -mercaptoetilamina ligada como uma amida ao cido pantotnico e tm um 4-fosfato. O 4-fosfato ligado 3`,5`adenosina difosfato por um pirofosfato na coenzima A. Na protena carreadora de acilas e na acil CoA sintetase, o 4-fosfato ligado a um resduo de serina. A forma utilizada na suplementao de dietas animais o pantotenato de clcio. 2 Absoro e metabolismo As formas coenzimticas do cido pantotnico so amplamente distribudas nos alimentos. Essas formas seriam hidrolisadas no intestino. O soro sanguneo possui predominantemente a forma livre do cido pantotnico. As enzimas celulares convertem o cido pantotnico em coenzimas. O excesso de pantotenato excretado via urina. 3 Funes da vitamina O cido pantotnico um componente da coenzima A, da acil CoA sintetase e da protena carreadora de acilas. Consequentemente, a forma coenzima A da vitamina, que se liga a grupos acila ativos, responsvel por reaes de condensao, trocas de grupos acila e transferncia de grupos acila catalizadas pelas respectivas enzimas. A coenzima A est envolvida tambm na degradao dos cidos graxos (atravs da formao de acil CoA). Os cidos graxos so sintetizados com a participao da protena carreadora de acilas. VITAMINA B6 (PIRIDOXINA) 1 As formas da vitamina A forma diettica predominante desta vitamina geralmente a piridoxina (PN), que a principal forma nos produtos de origem vegetal. O piridoxal (PL) e a piridoxamina so as principais formas encontradas nos tecidos animais. Todas as trs formas so convertidas no organismo animal nas formas metabolicamente ativas, o piridoxal fosfato
163
(PLP) e a piridoxaminafosfato (PMP). A forma sinttica de piridoxina utilizada para suplementao nas dietas geralmente o hidrocloreto de piridoxina (PN.HC1). 2 Absoro e metabolismo A microflora ruminal sintetiza a piridoxina em quantidades normalmente suficientes para satisfazer as necessidades dos poligstricos. A sntese microbiana corre tambm no clon dos monogstricos, sem que haja absoro em quantidade aprecivel. A absoro dessa vitamina hidrosolvel ocorre no intestino delgado por um processo passivo. Parece existir uma pequena estocagem orgnica de vitamina B6. Uma vez absorvida, a piridoxina convertida nas suas formas metabolicamente ativas, o piridoxal fosfato (PLP) e a piridoxamina fosfato (PMP). Essa converso requer a presena da flavina mononucleotdio (FMN), da flavina adenina dinucleotdio (FAD) e da nicotimamida adenina dinucleotdio (NAD). Consequentemente, uma deficincia de niacina ou de riboflavina, necessrias para sntese de NAD e FAD respectivamente, pode resultar em baixos nveis de formas ativas da piridoxina. O piridoxal fosfato atua com a quinureninase na sntese de niacina a partir do triptofano. 3 Funes As formas metabolicamente ativas da vitamina B6 so as coenzimas pirodoxal fosfato (PLP) e piridoxamina fosfato (PMP). O PLP est envolvido na maioria das reaes do metabolismo de aminocidos inclusive a transaminao, descarboxilao, dissulfidrilao e desaminao no oxidativa. O PLP participa ainda como cofator de uma enzima da sntese de porfirinas, e da fosforilase do glicognio (catabolismo do glicognio). Outra funo, no compreendida, est no metabolismo lipdico. O PLP importante no metabolismo do cido gama amino butrico no crebro e na sntese da adrenalina e noradrenalina a partir da fenilalanina ou da tirosina.
BIOTINA OU VITAMINA H 1 Absoro e metabolismo A biotina parece ser bem absorvida no intestino delgado, embora as formas ligadas a protena contidas nos alimentos no sejam prontamente disponveis para o animal. H evidncias de que a biotina no bem retida pelo organismo. A sua excreo, bem como a da maioria das vitaminas hidrossolveis est estreitamente relacionada com a ingesto. 2 Funes A biotina participa de reaes metablicas de carboxilao, como coenzima das respectivas carboxilases. As mais importantes das enzimas de carboxilao dependentes da biotina so a piruvato carboxilase, a acetil CoA carboxilase e a propionil CoA carboxilase. A primeira catalisa a converso do piruvato em oxalacetato. Esta reao importante na gliconeognese e na formao do glicerol. A propionil CoA
164
carboxilase por sua vez permite a sntese de succinil CoA, processo fundamental na gliconeognese (a partir do propionato) em ruminantes. A carboxilao da acetil CoA gera a malonil CoA, necessria para a sntese dos cidos graxos de cadeia longa. A biotina serve tambm para a fixao do carbono n 6 nas purinas, utilizadas na sntese de DNA e RNA. CIDO FLICO 1 Formas da vitamina O nome folacina uma designao genrica de todos os compostos que possuem a atividade biolgica do cido flico. O cido pteroil-glutmico foi a primeira forma de folato a ser isolada e sintetizada. Contudo, em quase todos os tecidos, as formas predominantes so os poliglutamatos. Essas formas provavelmente possuem at oito resduos adicionais de cido glutmico ligados entre si e ao glutamato terminal do cido flico por ligaes amida (poli--amida). Os poliglutamatos reduzidos so as formas coenzimticas ativas nos tecidos animais. O monoglutamato a forma de suplementao nas raes. O organismo animal converte o monoglutamato absorvido em poliglutamatos.
2 Funes O anel de pteridina do cido flico completamente oxidado e pode ser enzimaticamente reduzido a dihidro e tetrahidrofolato. As formas coenzimticas da vitamina so poliglutamatos na forma de tetrahidrofolatos. Elas carregam os grupos ativos monocarbonados metil, formil, formimino, metileno e metenil. Esses grupos derivam do formiato ou do metabolismo da glicina, serina e histidina e so transportados pelos tetrahidrofolatos e utilizados na sntese de compostos como a metionina, os aneis de purina bem como na converso da desoxiuridina monofosfato em desoxitimidina monofosfato. Conseqentemente as coenzimas flicas, o S-adenosil metionina e a vitamina B12 so responsveis pela transferncia de unidades monocarbonadas nas vias metablicas. REFERNCIAS BIBLIOGRFICAS ANDRIGUETTO, J.M., PERLY, L., MINARDI, I. et al. Nutrio animal. 1. As bases e os fundamentos da nutrio animal: Os alimentos. 4.ed. So Paulo: Nobel, 1990. p.135-172. LUDWIGSHAFEN, N.A., BONN, G.B., ELMSHORN, D.D. et al. Las vitaminas en la nutricin animal. Bonn: AWT, 1985. STRYER, L. Bioqumica. 4.ed. Rio de Janeiro: Guanabara Koogan, 1000p. 1996. SWENSON, M.J. Dukes / Fisiologia dos animais domsticos. 10.ed. Rio de Janeiro: Guanabara Koogan, 1988. p.383-395. VITAMIN tolerance of animals. Washington: National Academy of Sciences, 1987. 96p.
165
MINERAIS INTRODUO Os alimentos e a gua contm praticamente todos os elementos minerais conhecidos, nutricionalmente desejveis ou no, sob vrias formas qumicas, ou seja, em estado livre ou sob a forma de compostos orgnicos e inorgnicos, e em propores muito variveis. A alimentao mineral adquire uma maior importncia medida que submetemos os animais a uma maior presso de produo. A evoluo das caractersticas genticas, os sistemas cada vez intensivos de explorao e a utilizao de raes cada vez mais complexas, tornam necessrio um maior conhecimento das necessidades nutricionais pelos animais para se obter uma produo tima No existe qualquer classificao dos elementos minerais que possibilite uma explicao convincente do fato de que alguns so essenciais vida e outros no. De acordo com Lloyd et al. (1978), um elemento inorgnico (mineral) considerado essencial quando atende os seguintes critrios: - estar presente em concentraes aproximadamente constantes nos tecidos sadios de todos os animais, com pequenas variaes entre espcies; - deficincias em dietas adequadas em outros nutrientes resultam em aparecimento de anormalidades estrut0urais e/ou fisiolgicas reproduzveis; - a adio do elemento dieta deficiente evita ou recupera a anormalidade surgida; - as anormalidades induzidas por deficincias minerais devem ser acompanhadas por alteraes bioqumicas especficas, que so prevenidas ou revertidas ao estado normal, quando fornecemos o(s) mineral(is) no(s) nvel(is) adequado(s). Pelo menos 13 elementos inorgnicos so considerados essenciais, sendo estes: clcio (Ca), fsforo (P), potssio (K), sdio (Na), Cloro (Cl), magnsio (Mg), enxofre (S), zinco (Zn), ferro (Fe), mangans (Mn), cobre (Cu), iodo (I), selnio (Se). Alm destes, outros elementos como o vandio (V), cromo (Cr), nquel (Ni), estanho (Sn), molibdnio (Mo), silcio (Si), flor (F), arsnico (Ar), chumbo (Pb) e cobalto (Co) esto sendo investigados para avaliar se so essenciais ou no, devido ao fato de suas necessidades estarem cobertas pelas matrias primas da rao, por ser muito pequeno seu requerimento. Os minerais tm trs funes bsicas no organismo: 1. Estrutural: - Formao do esqueleto; Como componente de protenas e lipdios; Integridade da parede celular, mantendo a presso osmtica 2. Regulao dos Processos Orgnicos: - Clcio intervm na coagulao sangnea e contrao muscular; - Vandio regula a sntese do colesterol e fosfolipdios; - Ferro e cobre intervm na sntese da hemoglobina, etc. 3. Regulao do Metabolismo Energtico: - Intervm como cofatores que ajudaro na liberao da energia a partir dos alimentos na sua transformao em outros metablitos;
166
O Ca, P, Mg, Mn e V intervm na sntese e quebra dos enlaces de alta energia do ATP.
Os elementos minerais essenciais so classificados em dois grupos, macrominerais e microminerais, de acordo com as quantidades relativas que se exija na dieta. De um modo geral, os minerais possuem duas funes bsicas no organismo: - funo estrutural - como constituintes de ossos e dentes; - funo reguladora dos processos orgnicos - como a regulao do pH e equilbrio cido-base, manuteno da presso osmtica, tnus muscular, impulso nervoso, atividades enzimticas e hormonal, etc. Esta diviso se realiza segundo seus requerimentos totais, no quanto importncia. Por exemplo: o Ca (macromineral) necessrio para a osteognese, mas sem o Co (micromineral) o crescimento se paralisa como se houvesse uma deficincia total em Ca. Na prtica, no basta apenas realizar um aporte de minerais sem considerar os distintos fatores que vo influenciar sua absoro e, portanto, utilizao no organismo. Existem diversas circunstncias que vo influir, a nvel intestinal, na eficincia com a qual um mineral absorvido: 1 - Interaes entre minerais: a) Formao de precipitados insolveis quando dois ou mais ctions competem pelo mesmo nion. Este o caso do cido ftico, pois quando um sal solvel ionizado no intestino, o ction pode ser seqestrado por ele, formando fitatos, que so sais estveis e insolveis, o que os torna no absorvveis. Esta reao ocorre sobretudo com Ca, Zn e Fe. Por outro lado, pode ocorrer que quando a molcula ligante no esteja presente em excesso, a suplementao de um elemento pode aumentar a disponibilidade de outro ao reduzir-se suas possibilidades para formar complexos (p.ex., quando se adiciona Cu dieta a concentraes relativamente altas como estimulante de crescimento, aumentando-se a disponibilidade de elementos traos como o Zn, diminuindo a incidncia de paraqueratose). Tambm podemos citar o exemplo dos fitatos (hexafosfato de myo inositol) que ao ser analisado no laboratrio considerado como Ptotal, mas os monogstricos no so capazes de romper esta molcula por no existir suficiente fitase no intestino, tornando o P no assimilvel. Estes fenmenos de precipitao podem ocorrer tambm com fosfatos e oxalatos. b) Competio entre ctions pela mesma protena de transporte, para passar a parede intestinal. Um exemplo deste fenmeno ocorre entre o Fe e Cu, que so antagonistas, competindo pela transferrina (o Cu tem preferncia de unio, o que pode diminuir a absoro de Fe). c) Os processos enzimticos essenciais podem ser bloqueados pela troca de um cofator metlico por um metal inativo. Um exemplo o que ocorre com uma das enzimas essenciais sntese de porfirina (frao da hemoglobina) que ativada pelo Zn mas inibida pelo chumbo (Pb).
167
d) Quando um metal que forma parte de uma metaloenzima substitudo por outro, a atividade enzimtica pode bloquear, acelerar ou no variar. o caso da carboxipeptidase (uma metaloenzima de Zn). Quando Zn substitudo pelo Co h uma diminuio de atividade enzimtica. e) Quando h um aporte excessivo de um metal, no somente h uma menor absoro intestinal sendo que tambm h uma reexcreo no lmen intestinal do excesso de metal, o que pode acarretar excreo de outros metais durante o processo. f) Mesmo que tenhamos considerado estas interaes de forma isolada, geralmente se produzem simultnea ou consecutivamente mais de um processo. 2 Interaes Vitamina-Minerais: as vitaminas tambm podem interferir na absoro intestinal de minerais. Tal como o caso do aumento na absoro de Fe causado pela vitamina C, ou a necessidade de vitamina D para absoro do Ca atravs do intestino. Isto se complica mais se considerarmos as interaes entre vitaminas (p. ex., um excesso de niacina pode deprimir a vitamina D e interferir, portanto, na assimilao e uso do Ca). 3 Interaes entre Minerais e Gorduras: estas interaes podem influir na biodisponibilidade deste mineral no organismo. Um exemplo a interrelao existente entre os microminerais e os cidos graxos, formando sabes insolveis no trato digestivo. Alm disso, h interaes entre a sntese de cidos graxos essenciais e minerais. 4 Interaes Fibra-Minerais: diversos estudos tm demonstrado que a presena de fibra no digestvel interfere e diminui a absoro de grande parte dos minerais. Estes estudos tm se concentrado sobretudo na diminuio da absoro de Ca, P, Mg e Zn. 5 Interferncia pH-Minerais: o pH intestinal tem grande influncia sobre a absoro mineral j que, em geral, pHs alcalinos diminuem a absoro (exceto dos metais alcalinos) e os ctions tendem a formar precipitados insolveis quando o pH elevado. 6 Estimao das Necessidades: a meta da investigao dos microminerais relativa fisiologia da nutrio principalmente estabelecer dados de necessidade para os elementos individuais. Pelo fato da utilizao de um elemento estar influenciada por um grande nmero de fatores, til traar uma distino bsica entre necessidade lquida e bruta. A necessidade lquida resulta das perdas do corpo, principalmente com as fezes e urina, mas tambm com a transpirao cutnea e a partir dos produtos elaborados. A necessidade bruta inclui as quantidades dos elementos-traos os quais, tendo em conta o fator utilizao, se deve ingerir com a dieta para cobrir a necessidade lquida. H basicamente trs mtodos para medir a utilizao dos microminerais: os estudos de balano, a relao dose-resposta e o mtodo fatorial. a) Estudos de balano: neste estudo, o consumo do elemento-trao se compara com a excreo. Por razes prticas, se consideram somente a excreo por fezes e urina, enquanto por razes tcnicas, as perdas atravs da transpirao, pele e plos so ignoradas. Isto considerado para o animal adulto que no esteja em crescimento, gestao ou lactao. Para o organismo adulto, a nica concluso que se pode traar que se o balano negativo (excreo > ingesto), as necessidades no so cobertas. Para o organismo em crescimento ou reproduo, a necessidade no se cobre se o balano zero ou negativo.
168
b) Relao dose-resposta: quando a resposta de um critrio adequado se investiga como funo da dose, que pode variar de subtima a subtxica. Tais relaes se podem avaliar metodologicamente por anlises de regresso. Na figura acima podemos ver que a administrao tima quando as diferenas marginais caem abaixo de um valor definido ou voltam a zero. Os parmetros selecionados como critrio de resposta seriam especficos para um elemento e indicam sensivelmente as mudanas mais estreitas no status de administrao. A necessidade lquida de um mineral obtida mediante estudos dose-resposta pode mudar em funo da velocidade de crescimento. Isto significa que os requerimentos precisam ser deduzidos para cada tipo de crescimento e intensidade. c) Derivao fatorial das necessidades no mtodo fatorial para derivar necessidades (Kirchgessner et al., 1974 e Weigang & Kirchgessner, 1977/78) a necessidade lquida de manuteno (Nm) e de crescimento (Nc) se determinam: N=Nm + NC. A necessidade lquida para manuteno inclui perdas inevitveis, as quais so as qualidades liberadas por excreo e no reutilizao mais as quantidades perdidas na descamao celular, perdas de plo e sudorao. A necessidade lquida para o crescimento, se determina pelo tipo e intensidade do mesmo, atravs da reteno do mineral receptivo nos tecidos do animal. Este tipo de determinao, tem naturalmente suas limitaes, com respeito ao uso para humanos. Com a finalidade de derivar a necessidade bruta da necessidade lquida, se deve considerar a utilizao. MACROMINERAIS I - CLCIO 1 Distribuio Tissular O clcio no ocorre livremente na natureza, mas os seus compostos so extensamente distribudos, sendo o ction mais abundante no corpo animal. Aproximadamente 99% do clcio armazenado no corpo do animal est no esqueleto como constituinte dos ossos e dos dentes. Se apresenta em uma proporo aproximada de 2:1 com o fsforo inorgnico dos ossos. No sangue, o clcio encontra-se principalmente no plasma em trs estgios: - on livre (60%) - ligado a protena (35%) - misturado com cidos orgnicos como cido ctrico ou cidos inorgnicos como o fosfato (5%) - necessrio para a contrao muscular e a coagulao sangnea - um moderador da excitabilidade neuromuscular que est relacionada ao (K + quociente das concentraes sangneas dos ons, ( Ca + 2 ) + ( Mg + 2 )
169
2 Metabolismo Os fatores primrios que influenciam o metabolismo do clcio so o fsforo, a vitamina D, sistemas hormonais e a idade do animal. No animal em crescimento, ocorre uma reteno lquida de clcio no organismo. A quantidade que se armazena nos ossos e nos outros tecidos excede a quantidade que se perde nas fezes, urinas e suor. Nos adultos que no se encontram no perodo de gestao, lactao e postura, a quantidade de clcio ingerido iguala a quantidade que se perde; se so preenchidas as necessidades metablicas. 2.1 Absoro O clcio da dieta absorvido principalmente no duodeno e jejuno da maioria dos animais monogstricos. A absoro se efetua tanto por transporte ativo (gasto de energia) como por transporte passivo (difuso). Fatores que interferem na absoro e na necessidade de clcio A) Estado fisiolgico do animal A absoro de clcio maior em animais com alta necessidade de clcio; tais como animais em crescimento e fmeas gestantes. Em frangas, a absoro de clcio aumenta bruscamente entre o 10 e 14 dia antes do incio da postura devido a mudanas hormonais. PH a nvel intestinal os ctions tendem a formar precipitados insolveis quando o pH elevado, em funo da atividade da bilis na luz intestinal. Vitamina D O clcio atravessa a parede de natureza mucosa da clula duodenal, mas no consegue passar pela parede de natureza serosa. A absoro completa do clcio um procedimento ativo que se encontra sob o controle da protena fixadora de clcio (CaBP), a qual dependente da vitamina D. Antes da vitamina D atuar na absoro do clcio, h necessidade da ativao da mesma atravs de duas hidroxilaes. No fgado, o colecalciferol (vit. D3) sofre uma hidroxilao no carbono 25, formando 25-hidroxi-colecalciferol (25OH-D3); em seguida, no rim, sofre nova hidroxilao no carbono 1, formando 1,25-dihidroxi-colecalciferol (1,25-(OH)2-D3) que a forma ativa da vitamina D (figura 1). As enzimas responsveis pelas hidroxilaes so colecalciferol-25hidroxilase e 25-OH-D3-1-hidroxilase, respectivamente. Existem outros metablitos da forma ativa da vit. D, sendo o mais estudado o 24,25-(OH)2-D3, proveniente da hidroxilao do 25-OH-D3 pela enzima 25-OH-D3-24-hidroxilase. Existe algumas evidncias que este metablito necessrio para uma eclodibilidade normal dos ovos. Este mesmo sistema enzimtico pode promover uma hidroxilao no carbono 24 da forma 1,25-(OH)2-D3 resultando no 1,24,25(OH)3-D3, o qual apresenta apenas 60% da atividade da 1,25-(OH)2-D3.
B) C)
170
D) E)
F) G)
H)
Gordura diettica Altos nveis de gordura diettica, sobretudo saturada, aumenta a formao de sabes insolveis de cidos graxos com clcio. Disponibilidade biolgica nos alimentos A disponibilidade biolgica ou bidisponibilidade refere-se aquela poro do clcio (e de outros nutrientes) que efetivamente utilizada pelo animal, sendo expressa em porcentagem do total contido no alimento. H grande variao da biodisponibilidade do clcio dos alimentos, dependendo, principalmente, da combinao qumica ou da associao fsica com outros componentes, formando, em geral, quelatos de baixa solubilidade ou baixa digestibilidade. Podemos citar como exemplo os fitatos, fosfatos e oxalatos. O cido ftico, presente nos alimentos vegetais, um ester cido do cido hexafosfrico com o inositol (figura 2), ocorrendo sob a forma de sais. O Ca+2, Mg+2 e Zn+2 podem ligar-se muito fortemente nos grupos fosfatos do cido ftico formando fitatos. Os fitatos caracterizam-se como sais estveis e insolveis. O cido ftico presente no trigo, cevada e aveia mais ativo que o do milho, sorgo e sementes oleaginosas. Lactose, fsforo e magnsio Ao contrrio do fsforo e do magnsio, a lactose facilita a formao de um complexo lactose clcio que favorece a absoro de clcio por fenmeno de interao. Fibra de baixa digestibilidade A presena de fibra no digestvel interfere e diminui a absoro de grande parte dos minerais, sobretudo na absoro de clcio, magnsio e zinco. A fibra poderia formar uma matriz em torno dos nutrientes, dificultando assim, a absoro dos mesmos. Natureza do clcio diettico As fontes inorgnicas (p. ex. carbonato de clcio) so em geral melhor absorvidos que as fontes vegetais.
2.2 Mecanismo homeosttico A concentrao relativamente constante do clcio no plasma obtida mediante controle endocrinolgicos complexos. Quando observa-se uma hipocalcemia temos uma produo do hormnio da glndula paratireoide, o paratormnio (PTH), e um aumento na atividade da enzima 1hidroxilase a nvel de rim, intensificando, deste modo, a sntese de 1,25-(OH)2-D3. Acredita-se que o PTH alm de mobilizar o clcio dos ossos e aumentar a reabsoro renal de clcio, estimula o sistema renal da 1hidroxilase. A forma ativa da vit. D (1,25-(OH)2-D3) alm de promover o aumento na absoro de clcio a nvel intestinal, aparece ligada receptores na glndula paratireide que estimulam a liberao do PTH. Todos estes mecanismos e interaes visam elevar o nvel de clcio at atingir o padro normal (normocalcemia). O aumento da concentrao plasmtica de clcio acima dos valores normais (hipercalcemia), ativa a glndula tireide a liberar calcitonina (CT) hormnio produzido pelas clulas C ou parafoliculares da glndula. A CT diminui o nvel plasmtico de clcio inibindo a atividade do PTH sobre os ossos, ou seja, inibindo a mobilizao de clcio dos ossos. Quando o organismo apresenta normocalcemia ou hipercalcemia observa-se uma diminuio na atividade da 1-hidroxilase e um aumento na atividade da 24hidroxilase. O resultado destas alteraes o aumento da 24,25-(OH)2-D3 e a diminuio da 1,25-(OH)2-D3 circulantes na corrente sangnea. A dinmica do mecanismo homeosttico do clcio pode ser assim resumido:
171
Decresce o nvel plasmtico de Ca aumenta secreo de PTH aumenta reabsoro de Ca sseo; ao mesmo tempo, aumenta a sntese de 1,25-(OH)2-D3 aumenta sntese de CaBP aumenta a absoro intestinal de Ca normalizase o nvel plasmtico de Ca. Eleva-se o nvel plasmtico de Ca aumenta a secreo de CT cessa a reabsoro ssea normaliza-se o nvel de Ca no plasma.
2.3 Ossificao A ossificao do esqueleto requer que os produtos dos ons clcio (Ca+2) e fsforo (PO 4 ) se encontre no lquido que rodeia a matriz ssea e excedam o nvel crtico mnimo. Estes produtos esto presentes no tecido sseo de ossos e dentes sob a forma de hidroxiapatita de frmula geral Ca10 (PO4)6 (OH)2. A substncia interticial do tecido sseo formada por um sistema bifsico, no qual a fase mineral encontra-se envolvida pela matriz orgnica. A fase mineral consiste principalmente de ons clcio, fosfato e hidroxila, alm do on carbonato, dispostos sob a forma de cristais. Podem existir outros ons em menor quantidade depositados na matriz orgnica, tais como, flor, ferro, magnsio, mangans e alumnio. A estrutura da hidroxiapatita pode sofrer modificaes em decorrncia de substituies atmicas. Apesar de ligeiras substituies, a relao Ca:P tende a se manter aproximadamente no limite de 2:1. 2.4 Excreo As trs principais vias de excreo so as fezes, a urina e o suor (figura 3). A excreo fecal inclui a frao que no se absorve e a frao endgena, originria principalmente das secrees da mucosa intestinal, 20 a 30% do clcio fecal total. Na maioria das espcies, a excreo urinria geralmente menor que a excreo fecal. Aproximadamente a metade do clcio plasmtico, principalmente o clcio inico, se filtra no rim em condies normais, entretanto, mais de 99% absorvido. A perda de clcio pelo suor pouco significativa, com exceo no homem e no cavalo, nos quais a sudorao abundante. 3 - Funes As principais funes do clcio so: - Formao dos ossos, dentes e casca do ovo; - Manuteno do equilbrio cido-base; - Participao no sistema de coagulao do sangue; - Controle da excitabilidade dos nervos e msculos. 4 - Deficincia Uma carncia em Ca acarreta o raquitismo no animal jovem e a osteomalcia no adulto (a matriz do osso normal mas a mineralizao insuficiente); diminuio da produo de leite e ovos e aumento da incidncia de ovos quebrados.
172
A dieta rica em fsforo provoca a diminuio da absoro de clcio e consequentemente, o quadro de focinho torcido em sunos. Estas afeces so raras na atualidade: no se deve confundir com a osteoporose que se caracteriza por um suporte sseo insuficiente e cuja origem no est necessariamente relacionada com a nutrio mineral (carncia em protenas). 5 - Excesso Ingesto de quantidades de clcio acima das necessidades metablicas pode produzir: - Calcificao dos tecidos moles; - Interferncia na absoro e utilizao de outros minerais, como Mg, Mn, Fe, Cu e I; - Paraqueratose em sunos resultante de uma dieta rica em clcio, o que dificulta a absoro de zinco. O problema pode ser corrigido com a suplementao de cobre. O aumento da concentrao de cobre na dieta incrementa a disponibilidade de zinco, diminuindo a incidncia de paraqueratose; - Urolitase em frangas com mais de 18 semanas de idade; - Problemas de patas em frangos, principalmente quando se usam nveis baixos de fsforo. 6 - Fontes e Exigncia Nutricional A maioria das matrias primas vegetais utilizadas na fabricao de raes aportam baixos nveis de clcio, e muitas vezes, mesmo apresentando nveis considerveis de clcio, estes no so considerados disponveis, como no caso da farinha de soja, que possui 1,3% de clcio, mas somente 0,3% disponvel. Por isso, as necessidades de clcio em animais monogstricos, devem ser cobertas com a suplementao de clcio a fim de corrigir as deficincias deste mineral nas matrias primas de origem vegetal. As fontes de clcio mais utilizadas para a suplementao das dietas so: calcrio, farinha de ostra, fosfato biclcico, farinha de carne e ossos. A exigncia nutricional estimada em clcio disponvel, para os diferentes estgios fisiolgicos dos sunos e aves, podem ser obtidas nas tabelas brasileiras publicadas pela Universidade Federal de Viosa e para as demais espcies monogstricas nas tabelas do INRA, NRC e IRC. II - FSFORO O metabolismo do fsforo e do clcio esto intimamente relacionados. Ambos formam matriz inorgnica do esqueleto, e a absoro de cada um deles est ligada quantidade do outro presente na rao. 1 - Distribuio Tissular Cerca de 80% do fsforo se encontra no esqueleto como parte do cristal de hidroxiapatita, e os 20% restantes nos tecidos moles e lquidos corporais, se encontram em sua maioria, sob a forma inorgnica. No soro sangneo, o fsforo se encontra sob a forma inorgnica e orgnica, sendo a ltima, um constituinte dos lipdeos. Aproximadamente 10% do fsforo
173
inorgnico e encontra ligado s protenas sricas e dele, 50 a 60% apresenta-se ionizado. O fsforo nos diversos tecidos do organismo apresenta-se sempre em estado dinmico, em constante intercmbio entre os variados rgos. Ao contrrio do clcio, que para manter a homeostase sangnea mobilizado dos ossos e reabsorvido nos rins, o nvel de fsforo no plasma varia com a ingesto. Por isso, a deficincia de fsforo se nota mais rapidamente que a deficincia de clcio. 2 - Metabolismo O metabolismo do fsforo se pode estudar em termos de metabolismo sseo, metabolismo de fosfolipdeos e de compostos de alta energia de fosfatos tais como ATP, ADP, AMP e fosfato de creatina. 2.1 - Absoro A absoro do fsforo no tubo gastrointestinal se realiza atravs do transporte ativo e difuso passiva. O clcio e absorve principalmente na primeira metade do intestino delgado, enquanto a absoro de fsforo continua at a altura do leo. Consequentemente, h de se considerar, que a relao clcio/fsforo, varia segundo os diferentes stios de absoro. Fatores que afetam a absoro e a utilizao de fsforo A) Fonte de fsforo - A maior parte do fsforo contido nos vegetais encontra-se combinada com fitatos ou com inositol; o cido ftico contm seis molculas de fsforo para cada molcula de inositol (figura 2). A capacidade dos animais em utilizar o fsforo ftico depende da espcie, sendo os mais eficientes os bovinos e outros ruminantes, seguidos pelos no ruminantes, na seguinte ordem: eqinos, sunos e aves. Esta capacidade decorre da presena de fitase dos prprios alimentos ou dos microrganismos do trato gastrointestinal. Desta maneira, as necessidades dos animais monogstricos se expressam em fsforo disponvel e no em fsforo total, como em ruminantes. B) Relao clcio/fsforo - A absoro de fsforo se relaciona diretamente com a concentrao diettica do clcio. O excesso de fsforo em relao ao clcio, diminui a absoro de ambos e a hipercalcemia diminui a reabsoro de fsforo no tbulo proximal do rim. Ambos os casos decorrem da formao de cristais de fosfato de clcio insolveis ou de baixa permeabilidade. Entretanto, na hipocalcemia observase o efeito fosfatrico do PTH a nvel do rim, ou seja, inibe a reabsoro renal de fosfato. C) Vitamina D - A vitamina D auxilia a absoro intestinal de fsforo, regula sua homeostase e a mineralizao ssea. Nveis muito baixos desta vitamina resultam em irritabilidade muscular, tetania e mineralizao ssea imperfeita. D) Idade dos animais - Os animais adultos utilizam mais eficientemente o fsforo de origem vegetal do que os jovens. E) Acidose metablica - No quadro de acidose metablica (causada por cetose; diabetes millitus em que h produo de cidos -hidroxibutrico e acetoactico; acidose renal na qual ocorre falha na reabsoro de bicarbonato e este perde-se na urina; diarria, na qual o suco pancretico, que contm bicarbonato, no reabsorvido e perde-se; etc) a concentrao de bicarbonato (HCO-3) diminui. Isso
174
requer ao corretiva renal, ou seja, excreo de H+ e restaurao da concentrao de bicarbonato no plasma (figura 4). A secreo de H+ na urina resulta na acidez da mesma, porm a quantidade de cido que pode ser eliminada sob a forma de ons hidrognio livres limitada. A urina pode ser acidificada apenas at um pH em torno de 4,5. Isso significa que a maioria dos H+ excretados deve estar ligada por bases, dentre as quais as mais importantes so HPO 4 e amnia, formando H2PO4- e amnio, resepectivamente. Os ctions que ficam em quilbrio eltrico com HPO 4 no filtrado glomerular (a maioria dos ons sdio) substituda pelos H+ secretados e, assim, retorna para o sangue. Consequentemente, a relao Na:Cl no lquido tubular diminui, e a necessidade de fsforo aumenta em funo da maior perda na urina na forma de H2PO4-. F) Ferro, alumnio e magnsio - Altas ingestes de Fe, Al e Mg interfere na absoro do fsforo devido a formao de fosfatos insolveis. 2.2 - Mecanismo Homeosttico A homeostase sangnea do fsforo mais complexa do que a do clcio. O fsforo se encontra em equilbrio no somente com o fsforo sseo, mas tambm e expressivamente, com vrios compostos do fsforo orgnico. A excreo renal do fsforo est suficientemente controlada pela secreo do hormnio da paratireide e pelo metablito mais ativo da vitamina D (1,25-(OH)2-D3). 2.3 - Excreo O fsforo, igualmente ao clcio, secretado (fsforo fecal endgeno) e excretado via luz intestinal. Entretanto, esta perda no representa uma proporo to significativa como a excreo fecal de clcio. A maior parte da excreo de fsforo se faz pelos rins e este parece ser o principal regulador da concentrao sangnea de fsforo. O hormnio paratireoideo e o 1,25-(OH)2-D3 so os protagonistas desse mecanismo de excreo. Quando a absoro intestinal de fsforo baixa, o fsforo urinrio descende a um nvel mnimo e sua absoro nos tbulos renais chega a 99%. Por outro lado, a excreo total pode aumentar consideravelmente, quando o consumo de clcio proporcionalmente elevado, e a excreo total de clcio (urina e fezes) aumenta ante a deficincia relativa de fsforo. Este processo preserva a ossificao normal dos ossos que exige a relao clcio/fsforo = 2:1. 3 - Funes Atravs das formas orgnica e inorgnica, o fsforo participa no metabolismo animal com as seguintes funes: - componente estrutural dos ossos, dentes e casca de ovo; - componente de fosfolipdeos com papel importante no transporte de gordura e permeabilidade da membrana celular. Como tal, o fsforo se encontra presente praticamente em todas as clulas; - intervm no equilbrio cido-base, atuando como tampo de fluidos intracelular; - componente essencial de certas molculas intimamente relacionadas com:
175
. metabolismo energtico - fosfatocreatina, adenosina monofosfato (AMP), adenosina difosfato (ADP) e adenosina trifosfato (ATP); . sntese protica - cido desoxirribonucleico (ADN) e cido ribonucleico (ARN); . sistema enzimtico - carboxilas e flavoprotenas. 4 - Deficincia O sinal mais comum da deficincia de fsforo nos animais o raquitismo, acompanhado com freqncia de uma alterao do apetite, o qual leva o animal a mastigar madeira, tijolos e etc. Esta perverso do apetite deonimina-se popularmente de pica. medida que a deficincia progride e o apetite diminui, o crescimento do animal e a produo ficam comprometidos. Em aves, os animais deficientes podem morrer, especialmente em situaes de estresse por excesso de calor. As porcas podem apresentar problemas de fertilidade com estros silenciosos e dificuldade no parto. A relao clcio/fsforo muito importante, sobretudo em monogstricos. Quando a relao demasiadamente alta, observa-se o aumento da incidncia de problemas de patas em aves e aprumo em sunos. 5 - Excesso Desde o ponto de vista prtico, o excesso de fsforo antieconmico devido ao alto custo das matrias primas, fontes deste elemento. Dietas ricas em fosfato que mantm a relao Ca:P acima de 1:2, interfere na absoro de clcio e diminui a concentrao de clcio plasmtico. O organismo a fim de manter o nvel normal de clcio plasmtico induz o hiperparatireoidismo secundrio nutricional, que se manifesta em conseqncia da reabsoro ssea excessiva. A desmineralizao ssea produz osteodisrrofia fibrosa (substituio do tecido sseo por tecido conjuntivo) nos animais, independentemente da faixa etria. Dependendo do grau de gravidade da distrofia ssea, o animal pode apresentar manqueira e fratura espontnea de ossos longos. O fsforo quando ministrado em quantidade acima das necessidades, apresenta efeito laxante. A diarria acompanhada da perda fecal de fsforo e outros nutrientes, o que debilita muito os animais principalmente os mais jovens. O excesso de fsforo tambm interfere na absoro de clcio, magnsio e outros minerais. Os sunos em crescimento que padecem de uma absoro ssea severa produzida pelo consumo de dieta pobre em clcio e rica em fsforo, se sufocam devido o ablandamento das costelas, at o ponto que se inibem os movimentos respiratrios. Em aves, tanto a deficincia como o excesso de fsforo, pode provocar problema de anormalidades esquelticas e reduo na produo e na qualidade da casca do ovo. 6 - Fonte e Exigncia Nutricional O fsforo pode ser de origem orgnica ou inorgnica. Na formulao de raes, considera-se fsforo o componente mineral de maior custo. Somente 20 a 45% do total
176
de fsforo presente nos vegetais assimilvel pelos animais monogstricos. A porcentagem pode aumentar para 50% se a rao for destinada a animais adultos. Quando a disponibilidade biolgica do fsforo elevada, pode se diminuir o seu contedo do alimento. Nas matrias primas de origem animal, a utilizao de fsforo pelo animal aumenta de 90 a 100%. A dieta complementa-se com fosfato biclcico, fosfato monoclcico e outros fosfatos utilizados como fonte de fsforo. No Brasil, existe grande potencial de rochas fosfricas, aproximadamente 2,9 bilhes de toneladas, que tem promovido um crescente interesse como fonte alternativa de fsforo em raes de sunos e aves. As exigncias de fsforo nos vrios estgios de desenvolvimento podem ser obtidos nas mesmas fontes citadas para o clcio. 7 - Mtodos para Melhorar a Utilizao do Fsforo Ftico: Pode-se aumenta-la mediante fitases de fungos e bactrias, como no caso de Aspegillus, melhorando-se desta forma a utilizao do Pftico (Nelson et al., 1978). A melhora na utilizao do Pftico em gros midos pode ser devida a uma menor proporo do mesmo no gro imaturo, ativao de fitases naturais em gros midos ou fitases de origem microbiana produzidas por fermentao. Ao contrrio do que acontece com o Pftico, a maioria das fontes de Pinorgnico, tem uma concentrao elevada de Passimilvel. Os fosfatos solveis em gua (monoclcio, tripolifosfato sdicos) so muito boas fontes de P, mas seu preo limita o emprego nas raes. A solubilidade em gua dos fosfatos no uma condio necessria para a assimilao do fsforo. Existem fosfatos completamente insolveis que podem ser muito assimilveis. Pelo contrrio, indispensvel que o P seja extrado e posto em meio cido, ou seja, em condies similares s do tubo digestivo. 8 - Relao Clcio - Fsforo: em leites e animais em engorda, uma relao Ca:P acima de 1,3:1 na dieta, acarretou depresso do crescimento e do desenvolvimento sseo, enquanto que ao fornecer uma concentrao alta de Ca, no se apresentaram efeitos adversos mesmo chegando-se a uma relao 2,0:1. No que diz respeito relao Ca:P, pode haver influncia do fato que os dois minerais no so absorvidos no mesmo stio do trato gastrointestinal. O Ca se absorve principalmente na primeira metade do intestino delgado enquanto a absoro do P continua at a altura do leo. Devido a esta particularidade, a relao Ca:P, a nvel de absoro, pode no ser a mesma que a nvel de alimento. Tambm influi a velocidade de absoro relativa do Ca e P, dependendo esta, da solubilidade dos sais de Ca e P no intestino. Mesmo assim, o coeficiente de utilizao digestiva real (CUDr) diminui com a idade para ambos. III - MAGNSIO 1 - Distribuio tissular O magnsio se distribui amplamente em todo o organismo e depois do clcio e fsforo, se encontra no corpo em quantidades maiores que qualquer outro mineral. Os ossos contm 0,8% de Mg, correspondendo a cerca de 60 a 70% do magnsio presente no organismo. Os 30 a 40% restantes esto distribudos pelos tecidos moles e fluidos corporais. Nos tecidos moles, se encontra em maior concentrao no fgado e msculos esquelticos.
177
No sangue, est distribudo aproximadamente em torno de 75% nas clulas vermelhas e 25% no soro. 2 - Metabolismo O metabolismo do magnsio complexo e variado, e est muito relacionado com o metabolismo do clcio e fsforo. 2.1 - Absoro A absoro do magnsio no aparelho digestivo, se realiza principalmente no leo de forma passiva e sem relao alguma com a vitamina D. 2.2 - Mecanismo Homeosttico No se conhece claramente o controle homeosttico do magnsio no sangue e nos tecidos. Aparentemente, este controle feito pelo mecanismo de excreo. Aproximadamente 95% do magnsio nos rins reabsorvido, o restante eliminado atravs da excreo urinria. A frao fecal endgena excretada principalmente na poro proximal do intestino delgado, tal como o clcio. 3 - Funes As funes de maior expresso do magnsio so: - constituinte do osso, dente e casca do ovo; - ativao de processos enzimticos nos quais intervm o ATP No fluido intracelular que contm altas concentraes de Mg+ o ATP e o ADP existem principalmente como complexos MgATP-2 e MgADP-. Em muitas reaes enzimticas nos quais o ATP participa como doador de fosfato, sua forma ativa realmente o complexo MgATP-2 (figura 5). Assim, o magnsio ao formar complexos com o ATP, participa de diversas funes: contrao muscular, impulso nervoso, sntese de protena, lpides, cidos nuclicos e coenzimas, e etc. provvel tambm que haja um efeito do Mg sobre o msculo uterino, favorecendo o parto. 4 - Deficincia - Diminuio da produtividade - Disfuno muscular - Excitabilidade nervosa e aumento da mortalidade - Calcificao dos tecidos moles - Vasodilatao perifrica - Extremidades dbeis e retorcidas, hiperritabilidade, espasmos musculares, tetania e morte pode ocorrer em sunos quando a relao Ca:P maior que 1:2. 5 - Excesso O excesso de magnsio, provoca uma ao tranquilizante ou efeito depressor sobre o sistema nervoso central. Isto se deve a diminuio da liberao do acetil colina na unio neuromuscular e nos gnglios simpticos.
178
Outros efeitos considerveis so: - diminuio da palatabilidade; - efeito laxante com produo de fezes aquosas em aves; - aumento de ovos com casca quebrada; - Um ligeiro excesso reduziria os intercmbios sseos. 6 - Fontes e Exigncia Nutricional As fontes inorgnicas so melhor absorvidas que as fontes vegetais. A maioria das forragens e outras matrias primas utilizadas na formulao de raes, so ricas em magnsio. Raramente necessria a suplementao deste mineral. As fontes proticas, tanto a animal como a vegetal e os cereais so ricos em magnsio. Para o aporte extra de magnsio se utilizam fontes inorgnicas, tais como, o xido e o sulfato de magnsio. As exigncias nutricionais do magnsio para os diferentes estgios fisiolgicos das espcies monogstricas podem ser obtidas nas tabelas do NRC, do ARC e do INRA. IV - POTSSIO, SDIO E CLORO O potssio, sdio e cloro so estudados juntos por serem eletrlitos muito importantes na manuteno da presso osmtica dos lquidos extracelular e intracelular e na manuteno do balano cido-bsico. 1 - Distribuio Tissular Aproximadamente 90% do potssio corporal intracelular e se intercambia facilmente com o lquido extracelular. Por outro lado, o sdio se encontra principalmente no lquido extracelular, com menos de 10% no interior da clula. Aproximadamente metade do sdio intracelular se adsorve aos cristais de hdiroxiapatita nos ossos. O cloro atua com o bicarbonato para balancear eletricamente o sdio do lquido extracelular. Encontra-se quase que exclusivamente no lquido extracelular e como elemento essencial do suco gstrico. Resumindo, praticamente todas as clulas contm uma concentrao relativamente alta de potssio e uma concentrao baixa de sdio. O plasma sangneo e a maioria dos outros lquidos extracelulares, apresentam alta concentrao de sdio e baixa concentrao de potssio (figura 6). A parte esquerda de cada barra do grfico mostra a composio de ctions; a parte direita a composio de nions. As reas mais escuras representam a soma de componentes menores. Observe a grande diferena no contedo de sdio e potssio entre o plasma sangneo e o fluido intracelular. Estes gradientes so mantidos pela ATPase Na+ K+ da membrana plasmtica em praticamente todas as clulas do organismo. O gradiente de ons H+ no suco gstrico resulta da ao de uma ATPase transportadora de ons H+ nas clulas parietais do estmago. 2 - Metabolismo 2.1 - Absoro
179
Os ons Na+, K+ e Cl- no so absorvidos em quantidades considerveis no estmago. A absoro e d principalmente na poro superior do intestino delgado e em menor grau na poro inferior do intestino delgado e no intestino grosso. Os trs ons se absorvem tanto por procedimento ativo como passivo; dependendo do local de absoro. O sdio e o potssio atravessam as mucosas por transporte ativo no intestino e principalmente por difuso no estmago. O cloro se absorve por transporte ativo no estmago e na poro superior do intestino delgado, mas por difuso passiva no intestino delgado (poro inferior). 2.2 - Mecanismo Homeosttico O nvel plasmtico do sdio e potssio tende a estar controlado pela ao do hormnio aldosterona, um mineralocorticide produzido pela crtex da glndula suprarenal. Ele promove a reabsoro parcial do sdio nos tbulos renais e a perda de potssio pelos rins. A produo de aldosterona, por sua vez, encontra-se sob controle do hormnio adrenocorticotrfico da glndula pituitria anterior. Este hormnio de ao antidiurtica liberado em resposta a mudanas da presso osmtica do lquido extracelular induzido por privao de gua. Ao mesmo tempo, a liberao de aldosterona suspensa e consequentemente a reabsoro renal de sdio. Em situao inversa, quando a ingesto de gua elevada ou h uma diminuio do sdio plasmtico, ocorre a inibio do hormnio antidiurtico e a aldosterona passa a atuar na reabsoro urinria do sdio. O consumo de potssio geralmente excede vrias vezes o requerimento da necessidade metablica do animal. Ainda assim, a intoxicao por potssio no ocorre devido a capacidade do rim de controlar a sua excreo. A aldosterona tambm intervm na manuteno de potssio no organismo. Uma concentrao elevada de potssio no lquido extracelular estimula a secreo de aldosterona da mesma forma que a baixa concentrao de sdio plasmtico. Na deficincia de potssio, uma parte do sdio transferido ao interior da clula para substituir o potssio, e desta maneira manter o equilbrio osmtico e cido-bsico. A concentrao de on cloro no lquido extracelular tende a equilibrar-se em relao a concentrao de sdio no organismo. A excreo renal excessiva do on sdio eleva a concentrao do on bicarbonato (HCO3-) para que se escrete igual quantidade do on cloro (Cl-) atravs da urina. A relao entre esses trs ons fundamenta-se na manuteno da concentrao idntica de ctions e nions no plasma. A homeostasia do potssio e do cloro tambm est muito relacionada. A deficincia de um resulta na deficincia metablica do outro. A reabsoro de potssio nos tbulos renais, necessita da presena de cloro. Consequentemente, considera-se o cloreto de potssio mais efetivo do que qualquer outro sal de potssio, para compensar a deficincia de potssio. 3 - Funes 3.1 - Sdio e potssio Presso osmtica - O sdio extracelular e o potssio intracelular atuam atravs de uma bomba sdio-potssio na manuteno do equilbrio osmtico (figura 7). Como j foi apresentado, a concentrao de potssio no interior da clula bem maior que a sua concentrao no plasma sangneo, e de forma inversa ocorre com
180
o sdio. A manuteno desses gradientes atravs da membrana plasmtica depende do fornecimento da energia qumica do ATP. A membrana das clulas contm uma enzima denominada ATPase transportadora de Na+K+. Ela cataliza a hidrlise de ATP em ADP e fosfato orgnico e utiliza a energia liberada para bombear K+ para dentro e Na+ para fora da clula, contra o gradiente de concentrao. Para cada molcula de ATP hidrolizada, trs ons de Na+ so transportados para fora e dois ons de K+ para dentro da clula. A ATPase Na+ K+ das clulas do tbulo renal funciona permitindo uma perda constante de potssio na urina, enquanto a perda de sdio deve ser mantida em nveis muito baixos. Cofator enzimtico - O potssio atua como cofator enzimtico juntamente com o magnsio na transferncia do grupo fosfato do fosfoenolpiruvato para o ADP. A reao catalizada pela piruvatoquinase. Fosfoenolpiruvato + ADP Mg +2 K+ enolpiruvato + ATP
Transmisso do impulso nervoso - A bomba de Na+K+ tambm responsvel pela transmisso do impulso nervoso. Ela gera uma diferena de potencial eltrico na membrana axonal do neurnio, responsvel pela transmisso do impulso nervoso ao longo dos neurnios. Manuteno do equilbrio hdrico e do balano cido-bsico Componente de secrees digestivas Participao do transporte de nutrientes e outros eletrlitos atravs das membranas celulares. 3.2 - Cloro Atua no controle da presso osmtica extracelular e na manuteno do balano cido-bsico do organismo. secretado em grandes quantidades no estmago, para juntamente com o on H+ formarem HCl (suco gstrico). Ultimamente, a ateno dos investigadores voltou-se mais para determinar o efeito que o balano inico pode ter sobre os resultados de produo, do que nas quantidades absolutas dos ons a suplementar. Este aspecto especialmente importante quando se utilizam dietas complexas, onde a utilizao de soros lcteos, pesados ou substituio parcial da soja (rica em K+) por lisina sinttica, pode alterar o equilbrio. Tabela - Efeito do balano de eletrlitos sobre o rendimento de sunos* Na+ K+ Cl- meq/kg Variveis -85 0 100 175 277 Ganho mdio dirio (em kg) 0,57 0,65 0,66 0,69 0,67 Consumo x dirio** (kg) 1,27 1,45 1,43 1,48 1,49 Converso alimentar 2,23 2,22 2,18 2,15 2,21 *Adaptao de Patience et al., 1987 **Efeito linear (P<0,002) e quadrtico (P<0,04) do balano eletroltico da dieta. 4 - Deficincia
341 0,66 1,48 2,23
181
4.1 - Sdio Pobre aspecto da pele, plo e plumagem; Diminuio do apetite; Diminuio do ndice de converso e produo; Depravao do apetite (picacismo) - o animal lambe qualquer objeto e bebe a urina num intento de satisfazer sua necessidade; Canibalismo entre as aves.
4.2 - Potssio A deficincia de potssio rara em animais monogstricos. Os sinais m ais comuns se manifestam por: - Aumento da susceptibilidade ao estresse; - Depravao do apetite e anorexia; - Perda geral do tnus muscular - marcha instvel; - Atraso do crescimento, emagrecimento e morte.
182
4.3 - Cloro A deficincia de cloro, igualmente a deficincia de potssio dificilmente ocorre. Os principais sinais so: diminuio do ndice de crescimento e aparecimento de sintomas nervosos. 5 - Excesso Geralmente, a ingesto superior a que se necessita, traz como conseqncia a excreo rpida do mesmo pelos rins. Como conseguinte, pouco provvel que o sdio, o potssio e o cloro em excesso na rao possam protagonizar um quadro de intoxicao. Exceto se restringido o consumo de gua, limitando a excreo urinria ou quando o animal apresenta insuficincia renal. 5.1 - Sdio O NaCl se torna txico quando seu nvel est acima de 2% na rao. Os perigos so menores quando o animal dispe de gua a vontade. Nos ruminantes, o sal utilizado com xito para restringir consumo, entretanto, isto seria perigoso para os monogstricos, que so relativamente susceptveis a intoxicao com sal. Os sinais de intoxicao se manifestam por andar cambaleante paralisia e convulses. 5.2 - Potssio O excesso crnico de potssio ocasiona a hipertrofia da crtex suprarenal devido a necessidade de aumentar a produo de aldosterona. 5.3 - Cloro O excesso de cloro produz um aumento moderado do consumo de gua. 6 - Fontes e Exigncia Nutricional Devido ao alto contedo de potssio e do cloro na maioria das matrias primas utilizadas na alimentao animal, no freqente a adio suplementar destes ons. Todavia, as altas necessidades do animal do sdio em relao ao seu contedo nas matrias primas, faz necessrio sua suplementao em raes compostas. Em geral, todos os alimentos so ricos em cloro. As forragens apresentam altos teores de potssio e os gros baixos teores de sdio e potssio. As dietas para monogstricos so suplementadas dom sal comum para cobrir as necessidades de sdio. O nvel suplementrio de sal (40% de Na e 60% de Cl) na rao geralmente est em torno de 0,3 a 0,5% para a maioria das espcies monogstricas. Para aves poedeiras o sal da rao deve limitar-se a 0,23%, pois o aporte de cloro para estes animais no deve ultrapassar a 0,14%. O sdio que falta, pode-se ministrar sob a forma de bicarbonato, carbonato ou sulfato de sdio.
V - ENXOFRE
183
1 - Distribuio Tissular As protenas esto presentes em cada clula do corpo e os aminocidos que contm enxofre so componentes praticamente de todas as protenas. Assim, podemos afirmar que o enxofre se distribui de forma ampla em todo o organismo animal. 2 - Metabolismo As formas orgnicas do enxofre se absorvem facilmente atravs do transporte ativo que se realiza a partir da parte superior do intestino delgado. A absoro do sulfato inorgnico a partir do aparelho digestivo pouco eficiente. O sulfato inorgnico se excreta atravs das fezes e da urina. O enxofre que no se absorve, provavelmente se reduz na poro inferior do aparelho digestivo como sulfato. O enxofre endgeno fecal chega ao aparelho digestivo principalmente atravs da blis. O enxofre urinrio se encontra principalmente como enxofre inorgnico mas tambm como outros componentes orgnicos (tiossulfato, taurina, cistina, etc.). A maior parte do enxofre corporal se encontra nas protenas e desta maneira, a excreo urinria de enxofre tende estar associada a excreo urinria de nitrognio. 3 - Funes O enxofre atua principalmente como componente de compostos orgnicos, tais como: Vitaminas como a biotina e a tiamina. A biotina intervm no metabolismo dos lipdeos, carboidratos e protenas. A tiamina participa na utilizao eficiente de energia e no funcionamento timo do sistema nervoso. Aminocidos sulfurados como metionina,cistina e cistena. As aves cuja plumagem formado por uma grande quantidade de aminocidos sulfurados, apresentam necessidades maiores de enxofre do que os mamferos. Molcula de insulina e glutationa, importantes reguladores do metabolismo. Coenzima A.
O enxofre de natureza inorgnica se concentra fundamentalmente nas seguintes molculas: - Condroitina - uma mucoprotena com papel estrutural das cartilagens e tendes. Apresenta tambm um papel de interesse na integridade da pele e dos fenmenos de coagulao sangnea. - Heparina - anticoagulante atuante no sangue. - Sulfato - intervm na desintoxicao de certas substncias como indol e glicosdeos cianognicos. 4 - Deficincia Em geral, a deficincia de enxofre muito difcil de se provocar e diagnosticar. Se a carncia devida ao enxofre inorgnico, o animal utiliza enxofre orgnico protico a fim de compens-lo. Assim, os sinais mais evidentes da deficincia de enxofre anorexia e perda de peso. 5 - Excesso
184
A intoxicao por enxofre no um problema freqente, porque a absoro intestinal do enxofre inorgnico muito baixa. O elemento enxofre considerado um dos elementos menos txicos. A toxicidade depende da capacidade do animal formar H2S a partir de fontes de SO4 inorgnico, pois o H2S compete com o cianureto em sua toxicidade. 6 - Fontes e Exigncia Nutricional O enxofre inorgnico suprido atravs dos produtos de origem animal incorporados rao. Quantidades importantes de enxofre inorgnico e ministram como veculos, j que muitos microminerais so suplementados sob a forma de sulfato, tais como o sulfato de cobre, sulfato ferroso e sulfato de magnsio. O enxofre orgnico adicionado rao sob a forma de metionina, aminocido essencial para monogstricos. VI - REFERNCIAS BIBLIOGRFICAS DUKES, H.H. et al. Fisiologia dos Animais Domsticos. 10 ed. Rio de Janeiro: Guanabara Koogan, 1988. 799p. MINI-SIMPSIO DO COLGIO BRASILEIRO DE NUTRIO ANIMAL, 6, 1991, Campinas. Anais... Colgio Brasileiro de Nutrio Animal, 1991, 189p. SOARES JR., J.H. Calcium Metabolism and its control - a review. Poultry Science, v.63, p.2075-2083, 1984. VELOSO, J.A.F. Perspectivas de uso dos fosfatos de rocha nacionais na alimentao animal. Cad. Tec. Esc. Vet. Da UFMG, n.6, p.55-84, 1991. MICROMINERAIS Os oligoelementos esto presentes em quantidades muito pequenas nos tecidos (ppm) e atuam em doses nfimas. Ao contrrio dos macroelementos, no entram na estrutura dos tecidos, salvo em raras excees (iodo da tiroxina, ferro da hemoglobina). As carncias agudas com sintomas graves so raras na prtica, mas podem existir subcarncias com repercusso sobre os rendimentos zootcnicos. Tambm pode-se induzir carncias por excesso de outros fatores alimentcios, produzindo-se as interaes j descritas. Os mtodos de estimao das necessidades dos oligoelementos so geralmente muito globais e as variaes das concentraes nas raes so difceis de apreciar, de maneira que os aportes recomendados comportam inevitavelmente uma certa impreciso. I IODO (I) 1 Distribuio tissular O corpo animal contm menos de 0,0004% de iodo. Cerca de 70 a 80% do iodo corporal concentra-se na glndula tireide. Pode ser encontrado tambm no estmago, intestino delgado, glndulas salivares, pele, glndulas mamrias, ovrios e placentas. Existem diferenas a este respeito entre as diversas espcies.
185
2 Metabolismo 2.1 Absoro e armazenamento O iodo absorvido sob a forma de iodeto majoritariamente no intestino delgado e em pequenas quantidades no estmago. O iodo necessrio chega tireide atravs do sangue. Ele armazenado nos folculos tireoideanos, includos em uma protena coloidal da glndula. So utilizados para iodar resduos de tirosina na tiroglobulina, uma glicoprotena. 2.2. Biossntese de compostos tireideos Os compostos tiroxina (T4) e triiodotironina (T3) so formados a partir de L3 Monoidotirosina (MIT) e 3-3 Diiodotironina (DIT) que se originam da iodao da tiroglobulina. A forma T3 mais ativa do que a T4, mas encontrada em menor quantidade no sangue. 2.3. Transporte Praticamente todo o T3 e T4 recm-formados so transportados no plasma sangneos ligados a uma globulina (TLG tiroxina ligada a globulina) ou a uma pralbumina. Os nveis de TLG geralmente se encontram mais elevados nos animais jovens e durante a prenhez. Em um determinado momento, os hormnios tireoideanos, tiroxina e triiodotironina so liberados gradativamente no sangue pela ao de enzimas proteolticas especficas. Livres, eles so transportados s clulas alvo onde se ligam s protenas receptoras especficas que direcionam o T3 ou o T4 ao ncleo celular. Em resposta a interao dos complexos tiroxina-receptor com genes especficos, as clulas alvo so induzidas a sintetizar grandes quantidades de certas enzimas e sistemas enzimticos. O principal resultado a estimulao da velocidade metablica basal do animal. 2.4. Degradao dos compostos tireoideanos e excreo Cerca de 80% dos hormnios tireoideanos que chegam aos tecidos se degradam por meio da subtrao do iodo no fgado, rim, e outros tecidos. O iodo liberado nas clulas pode recircular e ser reutilizado e os resduos de tirosina podem ser catalizados ou utilizados para sntese de protenas tissulares. O organismo perde os 20% dos hormnios tireideos restantes atravs da excreo biliar. O iodo inorgnico se excreta principalmente atravs dos rins e em menor quantidade atravs do suor e da blis. As glndulas salivares secretam grande quantidade de iodo, mas a maior parte se reabsorve no aparelho digestivo. 2.5. Homeostase sangnea dos compostos tireoideos. Os hormnios tireideos so sintetizados em resposta a sinais recebidos do hipotlamo que secreta o hormnio liberador da tireotrofina (TSH) em resposta a uma produo reduzida de tiroxina (retro-alimentao), o que estimula a hipfise anterior a secretar a tireotrofina no sangue.
186
A tireotrofina ao se ligar aos receptores nas clulas da glndula tireide, estimula-as a produzir os hormnios tireideos. Entretanto quando a concentrao da tiroxina e da triidotironina no sangue est elevada, atua como inibidora da secreo de TSH pelo hipotlamo e da tireotrofina pela hipfise. Ademais, a somatostatina secretada pelo hipotlamo e tambm pelo pncreas pode inibir a secreo de TSH. Portanto, a secreo ou a ao da tiroxina e triiodotironina so regulados por vrios hormnios. 3. Funes - Componentes da tiroxina e triiodotironina; hormnios reguladores do ndice de oxidao celular ou metabolismo basal. - Influe indiretamente nos processos de termoregulao, crescimento corporal e desenvolvimento da pele, plos e plumas. - Influe na atividade gonadal. 4. Sinais de deficincia e excesso 4.1 Deficincia A deficincia de iodo nos animais jovens se denomina cretinismo e nos adultos mixedema. A deficincia diettica de iodo diminui o ndice de metabolismo basal e consequentemente prejudica o crescimento e a atividade gonadal. Os problemas reprodutivos se associam a reabsoro fetal, natimortalidade, abortos, estro irregular, diminuio da libido, piora da qualidade do smen e problemas com a incubabilidade dos ovos. A pele dos animais apresenta-se seca e o plo quebradio. As aves produzem emplumagem anormal. Os sunos cujas mes so deficientes em iodo, podem apresentar alopcia e a pele seca e grossa. Os animais adultos podem apresentar hipertrofia da glndula (tambm chamada de bcio) e diminuio da produo lctea. Pode haver interao do iodo com outros minerais (Mn, Co, etc.) ou a presena de compostos antitireoideanos na dieta como as substncias bociognicas (que aumentam os requisitos de iodo). Estas substncias so o 5-vinil-2-oxazolidinetiona, o 3 butenil o 4 butenil derivados do isocianato, este presente em algumas sementes. Dentre os alimentos que contm bociognicos citam-se a famlia Brassica (couve, nabo, acelga, etc.), o amendoim e a soja.
4.2. Excesso O excesso de iodo no organismo pode resultar de uma dieta mal equilibrada, de desinfeces ou tratamento contra micose com fontes iodadas. Como resultado do excesso de iodo os animais apresentam: - lacrimejamento incontrolado; - salivao excessiva; - congesto respiratria; - diminuio do consumo de alimento - a suplementao excessiva de iodo a fmeas gestantes causa nascimento de animais com bcio, porque h um estmulo do iodo para o aumento de
187
tamanho da tireide do feto. Este fato j foi observado em eqinos, ratos, aves e pessoas. 5 Fonte e exigncias nutricionais O contedo em iodo dos vegetais maior nas reas litorneas em relao ao interior do pas. A alfafa e produtos de origem animal so boas fontes de iodo. A suplementao de iodo se faz principalmente com iodeto ou iodato de potssio entretanto o iodato de clcio e o iodeto de cobre tambm podem ser utilizados. O iodato de potssio mais estvel na mistura mineral do que o iodeto de potssio. Este quando em contato com o sulfato de cobre da mistura mineral, perde rapidamente o iodo que se volatiliza. As exigncias nutricionais do iodo para os diferentes estgios fisiolgicos das espcies monogstricas, podem ser obtidas nas tabelas brasileiras da Universidade Federal de Viosa e nas tabelas do Institut Natoinal de la Recherche Agronomique (INRA), National Research Council (NRC) e IRC. Deve-se entretanto, ademais das recomendaes, considerar os problemas da variao da qualidade dos ingredientes, processamentos e estocagem. II FERRO (Fe) 1 Distribuio tissular Cerca de 70% do ferro corporal encontra-se sob a forma heme, presente nas clulas vermelhas do sangue e na mioglobina do msculo. Aproximadamente 20% se armazena sob formas lbeis no fgado, bao, medula ssea e outros tecidos, onde se encontra disponvel para a formao de hemoglobina. Os 10% restantes se encontram nos tecidos, como parte da miosina e actomiosina muscular. Est presente ainda nas enzimas metaloporfirnicas (citocromo C, catalase e peroxidase) e metaloflavnicas (desidrogenase, redutase, xantino oxidase e NADH). 2 Metabolismo Apenas uma pequena frao de 5 a 10% do ferro na maioria dos alimentos de fato absorvida sob a forma ferrosa (Fe2). Absoro mais eficaz em condies cidas. A quantidade de ferro absorvida no estmago e principalmente no duodeno, mais expressiva do que no leo. A xantino-oxidase da mucosa intestinal cataliza converso do ferro do estado ferroso ao estado frrico, facilitando sua ligao transferrina, uma protena especfica responsvel pelo transporte do ferro atravs da corrente sangnea. O ferro presente na carne melhor absorvido do que o ferro presente nos cereais. As concentraes elevadas de fosfatos inorgnicos e cidos ftico nos cereais, diminuem a absoro de ferro ao formar sais insolveis. Tambm, concentraes elevadas de outros elementos traos, com o Zn, Mn, Cu e Cd, reduzem sua absoro . Presumivelmente, isto se deve competio pelos stios de enlace de protenas de transporte presentes na mucosa intestinal. O controle de absoro de ferro aparentemente depende da concentrao de ferro sob a forma de ferritina nas clulas da mucosa intestinal. Do ferro captado somente uma pequena poro se transfere para as clulas do sangue; a maior parte se retm nas clulas da mucosa e desta forma esse transferem para a clula sangnea conforme sua necessidade.
188
Nos tecidos, o ferro (forma ferrosa) est armazenado sob a forma de ferritina. Quando a capacidade de armazenamento excedida, o ferro acumulado como grnulos insolveis de hemosiderina dentro das mitocndrias de alguns tecidos. A ferritina considerada a forma solvel, e a hemosiderina a forma insolvel do ferro armazenado. H um constante intercmbio do ferro entre os tecidos e o plasma. A incorporao de ferro plasmtico (transferina) ferritina nas clulas hepticas depende de energia (ATP) e se relaciona com a reduo de Fe3+ da transferina a Fe2+ da ferritina. A liberao de Fe2+ da ferritina heptica ao plasma se efetua atravs do catalizador xantino oxidase. O organismo retm o ferro que absorve. O ferro liberado da degradao da hemoglobina reciclado e utilizado na sntese da nova hemoglobina.. O ferro excretado constitudo principalmente pelo ferro diettico que no se absorve, pelo ferro endgeno eliminado pela blis e pelo ferro das clulas intestinais descamadas. A excreo urinria ocorre somente quando a administrao do ferro parenteral superior a capacidade do plasma em liga-lo. 3 Transferncia placentria e mamria A eficcia da transferncia de ferro ao feto atravs da placenta diferente entre as espcies. O ferro se transfere ao feto atravs de um processo ativo, e sua concentrao na circulao fetal excede a concentrao plasmtica materna. A transferrina no atravessa a placenta. Inicialmente necessrio que o ferro se dissocie da transferrina no lado materno da placenta, para posteriormente associar-se com protena transportadora do lado fetal. medida que progride a gestao, mais e mais ferro se transfere ao feto. Ainda que os recm-nascidos de algumas espcies apresentem uma concentrao de ferro heptico relativamente elevado, como no saco vitelino das aves, o suno recm-nascido ao contrrio, propenso a apresentar deficincia em ferro. A concentrao de ferro no leite de todas as espcies baixa. A tentativa de aumentar esta concentrao no sangue da reprodutora, atravs da administrao de suplementos de ferro dieta ou atravs da aplicao de ferro parental infrutfera.
4 Funes O tomo de Fe est presente no centro do grupo ferro-porfirina ou grupo heme (Figura 2) de vrias protenas (hemoglobina, mioglobina, transferrina, ferretina, ovotransferrina, e lactotransferrina) e enzimas (catalase, peroxidase, citocromo C, desidrogenase, NADH, redutase e xantino oxidase) que participam do transporte de oxignio e de eltrons da cadeia respiratria. 5 Sinais de deficincia e excesso 5.1 Deficincia Exceto em animais jovens a carncia de Fe difcil de se produzir na prtica. O sinal mais freqente de deficincia de ferro a anemia microctica hipocrmica que se
189
caracteriza por palidez, anorexia, respirao dificultosa, crescimento retardado e baixa resistncia s doenas. A anemia por deficincia de ferro um problema comum que se observa nos casos de infestao massivas com parasitos internos ou sunos recm-nascidos, sujeitos a uma transferncia placentria e mamria ineficaz. As reservas em Fe do leito ao nascimento so muito pequenas (50 mg) e se esgotam muito rapidamente, j que o leite apresenta uma forte carncia neste elemento (1 mg/litro), comparativamente com a sua necessidade (7 mg/dia). O suno ao nascer apresenta 10g/dl de hemoglobina; uma concentrao normal para suas necessidades metablicas. Entretanto com seu crescimento extremamente rpido, apresentando s trs semanas, cinco vezes o seu peso ao nascer, ocorre diluio do armazenamento com instaurao de anemia e problemas de crescimento. Sabe-se tambm que o aporte de Fe da me passa com dificuldade ao leite, e a administrao de Fe ao leito no momento do nascimento permite evitar a anemia: a injeo de Fe dextrano prefervel suplementao por via oral, j que por esta via se estimula o crescimento bacteriano, em particular da E. coli. A injeo intramuscular de 150-200mg de ferro-dextrano aos 2 ou 3 dias de vida, mantm o nvel normal de hemoglobina at a terceira semana quando o consumo de alimento seco proporciona a quantidade suficiente de ferro. Atualmente se dispe do Fe quelado com aminocidos, aumentando a assimilao do Fe e diminuindo os problemas. Mesmo assim, ultimamente se est estudando o emprego de uma enzima chamada lactoferrina, para aumentar a absoro do Fe a nvel intestinal com duplo efeito: aumento da biodisponibilidade e menor presena de Fe no contedo intestinal disponvel para enterobactrias. Convm evitar as carncias em vitamina E susceptveis de provocar acidentes (hemorragias, choques mortais) no momento da injeo. 5.2 Excesso As diferentes espcies de animais domsticos so muito tolerantes ao excesso de ferro. A intoxio crnica de ferro produz diarria, diminuio da taxa de crescimento e da eficcia de utilizao do alimento. Em raes que incluem farinha de algodo rica em gossipol, o excesso de ferro pode ser benfico. O gossipol e o ferro formam um complexo no txico para o animal. 6 Fonte e exigncias nutricional O leite contm pouco ferro, mas as matrias-primas concentradas so boas fontes do elemento. Em geral, a suplementao no necessria, j que as fontes adicionais de clcio e fsforo costumam conter altas quantidades de ferro com disponibilidade aceitvel. Entretanto, na prtica, comum adicionar ferro s raes compostas para alimentao de sunos e aves. A causa mais provvel o seu baixo custo. Sulfato ferroso a fonte suplementar mais utilizada. As exigncias nutricionais podem ser obtidas nas mesmas fontes citadas paera o iodo. III ZINCO (Zn) 1 Distribuio tissular
190
Na epiderme e anexos (pele, plos, l e penas) est a maior parte do zinco presente no corpo, mas se encontra tambm em concentraes mais elevadas no fgado, osso, rim, msculo, pncreas, olho, prstata, placenta. A concentrao de zinco no sangue se divide entre as clulas e o plasma em uma relao 9:1. A maior parte do zinco plasmtico se liga s globulinas e a maioria do zinco encontrado nas hemcias est presente como componente da anidrase carbnica. 2 Metabolismo A absoro de zinco que se processa no intestino delgado corresponde de 5 a 40% do consumo. A absoro de zinco se encontra afetada de forma adversa por uma elevada concentrao diettica de clcio e presena de fitatos de gros utilizados na elaborao da rao. O cido ftico, grupos fosfatos mltiplos, ao ligar o zinco forma um composto insolvel no absorvvel, reduzindo a disponibilidade do mesmo. O agente quelante, Etileno Diamina Tetra Acetato ou EDTA, aumenta a disponibilidade de zinco ao competir com o fitato para formar o complexo EDTA Zn facilmente absorvvel. Por outro lado, a injeo de EDTA aumenta a excreo de zinco. A histidina e a cistina encontrados em grande quantidade em alimentos, tais como a soja e o milho, diminuem a absoro do elemento. O zinco armazenado na clula epitelial da mucosa intestinal, onde se encontra includo a uma metaloprotena de baixo peso molecular, denominada metalotionena. Ela sintetizada em resposta ao aumento da concentrao plasmtica de zinco no interior dos eritrcitos. A transferncia de zinco das clulas da mucosa intestinal ao plasma esta controlada pela metaloprotena. A produo de metalotionena se faz mediante a sntese de tionena, uma metalotionena livre de metais. Figura 3 O zinco no absorvido e o zinco endgeno so excretados principalmente pelas fezes. A outra via de excreo do zinco endgeno que assume grande importncia o suco pancretico. As perdas urinrias no so muito significativas. Quando a concentrao do zinco plasmtico excede s necessidades metablicas imediatas, promove-se a formao de compartimentos intra-celulares e a excreo endgena de zinco. De forma similar, o fgado ao produzir metaloprotena colabora com a homeostase do clcio. Um estado de estresse pode determinar o aumento da atividade de glicocorticides e consequentemente desencadear o aumento da sntese heptica de metalotionena. Como resultado, observa-se o armazenamento de zinco no fgado e uma diminuio concomitante de zinco plasmtico. A m absoro do Zn tem carter hereditrio: lethal trait A-46 (caracterstica letal A-46), conhecida por paraqueratose hereditria ou hipoplasia hereditria do timo, afeta o gado da raa holandesa e Black Pied Danish, j descrita no Brasil (Arq. Bras. Med. Vet. Zoot., v.38, n.6, p.871-878, 1986). 3 Funes O zinco cofator de numerosas metaloenzimas: - anidrase carbnica intervm no transporte de oxignio e dixido de carbono no glbulo vermelho.
191
- Carboxipeptidase A e B e ribonucleases so enzimas secretadas pelo pncreas no intestino delgado onde catalizam a hidrlise de peptdeos e cido ribonucleico presentes no alimento ingerido. - Desidrogenase cataliza reaes bioqumicas de oxi-reduo. Os pares de coenzimas NAD+ e FAD participam - DNA polimerase: responsvel pela iniciao e sntese de cadeias de DNA. E ainda: - Participam na sntese e metabolismo de protenas. - Participa no metabolismo dos carboidratos por ser um componente da insulina - Atua na formao do esperma. 4 Sinais de deficincia e excesso 4.1 Deficincia Os sinais mais caractersticos da deficincia de zinco so atraso do crescimento e anorexia. freqente encontrar hiperqueratinizao das clulas epiteliais. A paraqueratose em sunos pode ser acompanhada de leses. Os animais ao serem alimentados com concentrao apropriada de zinco na dieta, se recuperam dentro de duas a trs semanas. Quando o cobre est numa concentrao relativamente alta na dieta, atuando como estimulante do crescimento aumenta-se a disponibilidade de zinco; diminuindo-se a incidncia de paraqueratose. As aves apresentam dermatite, plumagem anormal e anomalias de patas (perose). A deficincia deste mineral acarreta efeitos drsticos sobre os rgos reprodutores masculinos; causando hipogonadismo e outras disfunes. No retardamento de cicatrizaes de feridas, pode-se pensar em deficincia de zinco. 4.2. Excesso Concentraes elevadas de zinco interferem na absoro de cobre e ferro no aparelho digestivo e consequentemente provoca um quadro de anemia. Gastrite e enterite podem resultar da ingesto excessiva deste elemento. Em poedeiras, xido de zinco na dosagem 10.000 a 20.000 ppm utilizado para provocar muda forada pois inibe o consumo. 5 Fontes e exigncias nutricionais As matrias primas mais ricas em zinco so os concentrados proticos e sobretudo as farinhas de carne. Os concentrados proticos vegetais no so boas fontes, j que o zinco apresenta-se quelado pelos fitatos. Para isto se utilizam fontes inorgnicas tais como: xido de zinco, sulfato de zinco e carbonato de zinco. As exigncias nutricionais do elemento so encontrados nas mesmas fontes citadas para o iodo. IV COBRE (Cu) 1 Distribuio tissular
192
Maiores concentraes de Cu ocorrem em animais mais jovens. Na maioria das espcies, as maiores concentraes de cobre se encontram no fgado, crebro, corao, rim, parte pigmentada do olho, plo, pncreas e msculo e em baixas concentraes na tireide, pituitria, timo, prstata, ovrio e testculo. O cobre no plasma se associa em cerca de 90% com a alfa 2-globulina como ceruloplasmina; apenas 10% se encontram nas clulas vermelhas do sangue como eritrocuprena. 2 Metabolismo O cobre se absorve no estmago e principalmente no intestino delgado. O pH do contedo intestinal modifica a absoro; sais de Ca diminuem a absoro de Cu ao elevar o pH. Os minerais Mercrio, Molibdnio, Cdmio e Zinco tambm prejudicam a absoro de Cu. O Hg e o Mo provavelmente pela formao de compostos insolveis com o cobre; o cdmio e o zinco por deslocarem o cobre ligado protena na mucosa intestinal. Algumas formas combinadas de cobre so melhor absorvidas do que outras. O nitrato cprico, o cloreto cprico e o carbonato cprico se absorvem melhor que o xido cuproso. O sulfato cprico mais facilmente absorvido que o sulfato cuproso. O cobre metlico apresenta baixa absoro. O cobre absorvido liga-se albumina plasmtica para ser transportado ao fgado, onde armazenado, e utilizado na sntese de inmeras protenas e enzimas, ou liberado ao plasma como albumina cprica e em grande quantidade como componente da ceruloplasmina. Na medula ssea, o cobre captado para formar a eritrocuprena das clulas vermelhas do sangue. O cobre excretado pelas fezes via blis. Atravs da clula intestinal e das secrees pancreticas se perdem quantidades menores, e pela urina quantidades insignificantes.
3 Funes: - Participa da hematopoiese ao permitir a absoro, o transporte e a liberao do ferro das clulas hepticas. - Componentes de enzimas metaloprotenas. A citocromo oxidase contm ferro e cobre no seu grupo prosttico. A lisil oxidase produz interligaes entre as cadeias polipeptdicas do colgeno e da elastina presentes nos vasos sangneos - Importante na formao de mielina e adrenalina . - Participa na formao dos ossos e no processo de pigmentao do plo e formao de plumas. - Favorece a absoro de zinco. - Mantm os processos de reproduo. - Intervm na formao de melanina e tem um papel importante na integridade da pele. 4 Sinais de deficincia e excesso 4.1 Deficincia
193
No muito freqente encontrar deficincias de cobre em animais criados intensivamente, devido: - riqueza de cobre nas matrias-primas. - s mnimas necessidades do animal - a capacidade de armazenamento no fgado A deficincia pode estar associada presena de fitatos, ao excesso de protena na rao ou interaes com outros minerais tais como zinco, ferro, molibdnio ou sulfato. Os sinais mais especficos de deficincia so: - anemia ferropnica (microctica normocrmica). - problemas cardiovasculares - problemas gastrointestinais - perda de rigidez dos ossos longos e aparecimento de ovos com casca disforme. - alteraes na pigmentao da pele, descolorao e perda de plo, emplumagem anormal, dermatite e comprometimento da qualidade da l. - ataxia com queda do tero posterior devido desmielinizao dos nervos. - falhas na reproduo com aumento da infertilidade e mortalidade fetal. 4.2 Excesso: Este elemento utilizado em doses elevadas em sunos (125 ppm), coelhos (100 ppm) e frangos (75 ppm) como estimulante do crescimento. Estas espcies so muito tolerantes a concentraes elevadas de cobre, o que no acontece com os ruminantes. Em frangos, o excesso de cobre aumenta as necessidades de aminocidos sulfurados e altera a integridade da moela, com aparecimento de lceras. O excesso de cobre em sunos pode aumentar a insaturao dos cidos graxos do toucinho, com incidncia de gordura blanda. 5 Fontes exigncias nutricionais As melhores fontes de cobre so os concentrados proteicos vegetais. Os cereais contm pequena quantidade de cobre, entretanto sua biodisponibilidade est em torno de 70%. A suplementao de cobre se faz com sulfato de cobre, xido de cobre ou carbonato de cobre. A disponibilidade do metal nestes componentes de 25, 80 e 53% respectivamente. As exigncias nutricionais podem ser obtidas nas mesmas fontes citadas para o iodo. V MANGANS (Mn) As necessidades de mangans na dieta dos animais, so bem menores comparando-as com as de outros minerais indispensveis. O contedo total de mangans em animais adultos somente 1% da quantidade total de zinco. Um homem de 70 kg, possui apenas 12-20 mg de Mn. 1 Distribuio tissular
194
O mangans se encontra disseminado por todo o corpo. Ao contrrio dos demais elementos traos, ele normalmente no se acumula no fgado e outros tecidos quando ingerido em quantidades elevadas. encontrado em maior concentrao no osso, rim, fgado, pncreas, glndula pituitria e crebro. Uma grande proporo de mangans nos tecidos moles se encontra sob a forma intracelular lbil, mas no osso, associa-se principalmente frao inorgnica. 2 Metabolismo A forma e controle da absoro do mangans no aparelho digestivo no so bem conhecidas. O processo pouco efetivo; geralmente a absoro menor do que 10%. Ele absorvido no aparelho digestivo como Mn, em seguida, sofre oxidao, formando Mn2+, depois nova oxidao, e como Mn3+ transportado pelo sangue ligado a uma betaglobulina plasmtica (transferrina). O mangans se concentra nos tecidos ricos em mitocndria (fgado, pncreas e crebro), todavia, se desconhece o papel exato deste elemento trao no metabolismo e sua funo na mitocndria. As funes so deduzidas atravs dos sinais de deficincia desenvolvidos pela carncia de mangans na dieta. A excreo do mangans se faz principalmente atravs da blis e em menor escala atravs do suco pancretico, urina e suor. Considerando-se que a homeostase corporal normal se mantm atravs da excreo e no da absoro. 3 Funes - O mangans indispensvel para a formao do sulfato de condroitina um componente da matriz orgnica do osso. - Essencial no metabolismo de todos os princpios nutritivos. - Mantm o funcionamento normal do sistema nervoso central e sistema reprodutivo. - Apresenta funo lipotrpica em sunos e poedeiras. - Componente da enzima arginase (Figura 4) os trs aminocidos esto relacionados sequencialmente. A ornitina precursora da citrulina, e esta, precursora da arginina. A arginina hidrolizada irreversivelmente a ornitina e uria pela ao da enzima arginase. Aps a regenerao, a ornitina participa de um novo ciclo da uria. - Ativador eficaz da enzima glicosil transferase responsvel pela sntese de mucopolissacrides e glicoprotenas. 4 Sinais de deficincia e excesso 4.1 Deficincia A deficincia em mangans pode estar acompanhada de anormalidades esquelticas irreversveis, relacionadas com a formao de ossos do corpo e do ouvido durante a vida pr-natal. Nos sunos jovens, observa-se encurtamento e arqueamento da pata, aumento das articulaes e ataxia. Os frangos jovens apresentam bico de papagaio, encurtamento e engrossamento das pernas e asas. As galinhas poedeiras respondem deficincia de
195
mangans com menor produo de ovos, e estes apresentam menor taxa de incubao e espessura da casca. O excesso de protena e a interao do mangans com outros minerais em grande quantidade na rao, acentuam a sua deficincia. 4.2 Excesso A intoxicao por excesso de mangans difcil de ser observada devido a grande tolerncia exibida pelos animais. Os efeitos de excesso de mangans se relacionam mais com a interferncia na utilizao de outros minerais do que com efeito especfico do prprio elemento. A utilizao de clcio e fsforo modificada quando o mangans encontra-se acima do requerimento normal. Em resposta, o animal diminui o consumo, apresenta menor ndice de crescimento e nos casos crnicos, raquitismo. Anemia por antagonismos com o ferro tambm pode ser detectada. 5. Fontes e exigncias nutricionais As melhores fontes de mangans so os derivados de trigo (farelo e farelinho) e as farinhas animais. O aporte adequado feito atravs de fontes inorgnicas, tais como: carbonato, xido e sulfato de mangans. Para os monogstricos, a forma sulfatada a mais utilizvel. As exigncias nutricionais de mangans podem ser obtidas nas mesmas fontes citadas para o iodo. VI SELNIO (Se) Por muitos anos o selnio, na forma de selenito ou selenato, foi considerado como txico para o gado criado em pastagens onde os sais de selnio esto naturalmente presentes em grande quantidade. Posteriormente descobriu-se que o selnio em pequena quantidade, necessrio na dieta de frango e sunos criados em confinamento. 1 Distribuio tissular O selnio encontra-se em todas as clulas do corpo. Concentraes mais elevadas so encontradas no fgado, rim e msculo. 2 Metabolismo O principal local de absoro do selnio no duodeno, onde se absorve de forma relativamente eficaz. A absoro aumenta em resposta s necessidades tissulares. A forma de administrao de selnio na dieta influencia significantemente a absoro. O selnio vegetal sob a forma inorgnica apresenta maior disponibilidade biolgica do que o encontrado nos tecidos animais, onde apresenta-se principalmente sob a forma orgnica. Depois da absoro, o plasma transporta o selnio em associao com uma protena plasmtica. Nos tecidos, ele armazenado principalmente como selenocistina
196
e selenometionina. A concentrao do elemento nos diversos rgos varia com o consumo. O selnio se incorpora s clulas vermelhas do sangue, leuccitos, mioglobina, nucleoprotenas, miosinas e vrias enzimas, que incluem o citocromo C e aldolase. A excreo responde aos requerimentos tissulares. As perdas se efetuam atravs dos pulmes (hlito com cheiro de alho), fezes e urina. As perdas fecais se constituem do selnio que no se absorve e do selnio excretado atravs da blis, do suco pancretico e das clulas da mucosa intestinal. 3 Funes As funes metablicas do selnio esto inteiramente relacionadas com as da vitamina E, e em muitas ocasies impossvel diferenci-las. Antioxidante, vitamina E e selnio tem funes semelhantes atuando para evitar a oxidao. Os antioxidantes dietticos evitam a formao de perxidos na rao. A vitamina E, um antioxidante biolgico, evita a formao de perxidos no organismo. O selnio destri os perxidos que conseguiram formar-se apesar da ao dos oxidantes e da vitamina E. Ao atuar corno componente da enzima glutationa-peroxidase, o selnio participa na reduo dos perxidos a lcoois de natureza no txica. O stio ativo da glutationa-peroxidase contm selenocistena; um aminocido raro, no qual o tomo de enxofre da cistena foi substitudo pelo tomo do selnio (Figura 5). Assim, o selnio como integrante da enzima gutationa peroxidase responsvel pela manuteno da integridade das membranas biolgicas contra a oxidao por perxidos intracelulares. E ainda, o selnio: - necessrio na formao de cistina a partir de metionina. - Preserva a integridade do pncreas, responsvel, atravs da lipase pancretica, pela absoro normal dos lipdios e dos tocoferis no aparelho digestivo. - Atua como cofator da aldolase, enzima que cataliza a clivagem da frutose 1,6-difosfato em duas trioses fosfatos: gliceraldedo 3-fosfato e diidroxiacetona fosfato. Este passo representa a ltima reao da primeira fase da gliclise. - Componente do citocromo C, protena mitocondrial transportadora de eltrons. 4 Sinais de deficincia e excesso 4.1 Deficincia muito difcil diferenciar os sintomas tpicos de uma deficincia em selnio dos sintomas de deficincia em vitamina E. Em geral, a vitamina E parece ter a capacidade de prevenir todos os sintomas de carncia do selnio. Na distrofia muscular nutricional (degenerao de Zenker) o msculo cardaco e s vezes o fgado, apresentam uma aparncia esbranquiada. A necrose heptica, igualmente distrofia muscular mais freqente em sunos jovens. A morte sbita pode resultar do comprometimento desses dois importantes rgos. Nas aves, a ditese exsudativa se caracteriza por acmulo de lquido subcutneo no peito, resultante do extravazamento da frao lquida do sangue aos espaos extracelulares. A encefalomalcea em frangos se cura somente com vitamina E ou com antioxidantes.
197
A vitamina E que tambm protege os animais contra a necrose heptica, ditese exsudativa e distrofia muscular, no tem efeito sobre as atividades da glutationa peroxidase por conseguinte, a ao protetora da vitamina E diferente da ao antioxidante do selnio. A carncia em selnio pode ainda ser responsvel por: - Reteno de placenta em ruminantes; - Maior incidncia de fgado gordo em poedeiras; - Menor resistncia a doenas. 4.2 - Excesso A toxidade por selnio no freqente em animais estabulados e sim em animais que pastoreiam em solo selenfero ou que se alimentam de vegetais cultivados neste tipo de solo. As dietas que contenham 5 ppm de selnio produzem sinais txicos na maioria das espcies. O consumo de dietas ricas em protena ou sulfato inorgnico, e a adio de arsnio reao ou a gua de beber, diminuem os sinais de intoxicao em sunos (manqueira, m formao e necrose dos cascos) e em aves (desenvolvimento embrionrio anormal e diminuio da produo de ovos e da taxa de incubao). 5 Fontes e Exigncias Nutricionais As protenas animais e a farinha de pescado so boas fontes selnio, entretanto sua disponibilidade baixa. Para o aporte inorgnico se utiliza selenito de sdio que apresenta considervel disponibilidade. As exigncias nutricionais podem ser obtidas nas mesmas fontes citada para o iodo. VII COBALTO (Co) 1 Distribuio Tissular Esta presente nos animais na quantidade de 1,1 mg. 43% est nos msculos, 14% nos ossos, 19%, na medula. Existe uma controvrsia a respeito do que sejam os padres normais e concentraes que indiquem deficincia (McNaugth: 0,04-0,06 ppm e 0,08-0,12 ppm; outros autores: 0,06-0,09 ppm e 0,28-0,34 ppm para concentraes de deficincia e concentraes normais respectivamente). 2 Metabolismo Acreditava-se que o cobalto fosse mal absorvido, mas atravs de pesquisas, concluiu-se que a sua absoro varia de 20-95% e que feita no intestino. Diminui a absoro quando o cobalto se liga protena e h um aumento quando a dieta pobre em ferro pois h indcios de que o stio de ligao desses dois minerais seja o mesmo e o mecanismo de regulao do transporte idntico. A principal via de excreo a urina, mas tambm excretado via fezes, suor e plos.
198
3 Funes - Representa aproximadamente 4,5% do peso da vitamina B12 e portanto est ligado hematopoiese e ao crescimento. - Aumenta a ao das peptidases. - Supe-se que diminua a ao de enzimas respiratrias como a citocromo oxidase e succinato-desidrogenase. 4 Sinais de Deficincia e Excesso Na deficincia os sintomas apresentados so: Anemia, anorexia e perda de peso, j quando ocorre excesso ocorre tambm anemia causada pelo impedimento da absoro do ferro, depresso do crescimento, anorexia e policitemia. 5 Fontes e Exigncias Nutricional Segue as mesmas recomendaes da bibliografia indicada para o iodo. VIII CROMO (Cr) 1 Distribuio Tissular Encontra-se principalmente no corao, aorta, pulmo, bao, rins e fgado. H uma diminuio do cromo com o avano da idade. Na aorta de pacientes com ocluso no foi encontrado cromo. 2 Metabolismo um mineral pobremente absorvido (apenas 1-3%). Sua absoro se d no intestino delgado (jejuno principalmente). O cromo um antagonista do zinco, e sua absoro aumentada pela presena de oxalatos. J os fitatos diminuem-na. Sua absoro maior na forma hexavalente. Esta ltima transportada pelas clulas vermelhas do sangue enquanto a sua forma trivalente transportada ligada betaglobulina. O transporte aos tecidos feito pela transferritina. A concentrao maior nos tecidos que no plasma. O cromo absorvido se excreta principalmente pela urina, mas tambm se encontra cromo nas fezes principalmente o exgeno porm h tambm cromo originrio da blis. 3 Funes um potencializador da insulina e portanto est ligado ao metabolismo da glicose. Estimula a sntese heptica de cidos graxos e colesterol diminuindo o colesterol circulante. 4 Sinais de Deficincia e Excesso A deficincia leva diminuio do crescimento e da longevidade, pode levar diabete mellitus e a arterioesclerose. O excesso causa depresso do crescimento e leses hepticas e renais.
199
As fontes e exigncias nutricionais so encontradas nas mesmas tabelas utilizadas para o iodo. IX FLOR (F) O flor assume importncia na nutrio animal quando se cogita a utilizao dos fosfatos de rocha ou fosfatos no defluorizados. Sabe-se que com o uso prolongado o flor alm de se acumular nos ossos acumula-se tambm nos tecidos moles. Nos ossos a formao da flUorhidroxiapatita confere-lhes maior rigidez mas a sua acumulao nos tecidos moles tem se mostrado malfica, apesar de alguns autores no aceitarem este fato. Sabe-se que facilmente absorvido no, intestino e excretado pela urina. Demais dados a respeito desse mineral so controversos e necessitam de maiores estudos. importante ressaltar que ele possui um efeito acumulativo e tambm se credita ao flor efeito cancergeno. X - MOLIBIDNIO (Mo) O molibidnio encontrado principalmente no fgado, ossos e rins. rapidamente absorvido no intestino. Sua excreo se d pela urina. Sabe-se que os sulfatos aumentam a excreo e diminuem o seu armazenamento. componente da xantino oxidase que responsvel pela formao do cido rico a partir da xantina. componente tambm da aldedo oxidase. Suas fontes e exigncias so encontradas nas mesmas referncias do iodo. XI MINERAIS QUELATADOS: Levando-se em considerao os fatores que influem na absoro dos diferentes minerais, fcil deduzir que independentemente do conhecimento da concentrao de um determinado elemento na dieta, muito difcil realizar uma predio do grau de absoro e equilbrio mineral, quando se utilizam fontes clssicas de aporte inorgnico destes oligoelementos. Uma forma de evitar-se esta situao a utilizao de minerais quelatados com aminocidos. Efetivamente, como um sal, o mineral est livre para participar das reaes descritas anteriormente, mas como um quelato estvel de aminocido, no h ionizao no intestino sendo, o mesmo, absorvido por via diferente. Um quelato se produz quando o mineral fixado a 2 aminocidos, formando um duplo anel heterocclico de baixo peso molecular. Nesta situao, no h precipitao do mineral dos sais nem h efeito da fibra ou gorduras sobre sua absoro. Igualmente, a utilizao de minerais quelatados com aminocidos oferece uma maior absoro, assim como a vantagem de no depender da situao do meio ambiente intestinal. Com relao a este tema, ultimamente a literatura especializada j bastante ampla, sendo inclusive realizados simpsios anuais onde so apresentadas s ltimas pesquisas sobre os produtos correlatos.
200
REFERENCIAS BIBLIOGRFICAS AMMERMAN,C.B. e GOODRICH, R.D. Advances in mineral nutrition in ruminants. J. Anim. Sci. 57(2) p.519-533, 1983. BREVES, G. e SCHRODER Comparative aspects of gastrointestinal phosphorus metabolism. Nutr. Res. Rev. 4, p.125-140, 1991. LEWIS, L.D. Alimentao e cuidados do cavalo. Ed. Roca. So Paulo, 248p. 1985. MATEOS, G.G. e BLAS, C. Minerals, vitamins and additives. IN: BLAS, C. e WISEMAN, J. The nutrition of the rabbit. Cabi publishing, Wallimgford, 1998, p.145-176. MCDOWELL, L.R. Mineral in animal and human nutrition. Academic Press: San Diego. 524p. 1992. MEYER, H. Alimentao de cavalos. Livraria Varela, So Paulo. 303p. 1995. NATIONAL RESEARCH COUNCIL. Nutrient requirements of horses. 5 ed. National Academic Press, Washington, 1989, 91p. NATIONAL RESEARCH COUNCIL. Nutrient requirements of poultry. 9 ed. National Academic Press, Washington, 1994, 155p. NATIONAL RESEARCH COUNCIL. Nutrient requirements of swine. 9 ed, National Academic Press, Washington, 1998, 93p. NUNES, I.J. Nutrio animal bsica. Belo Horizonte, Ilto Jose Nunes, 334p. 1995 PEELER, H.T. Biological availability of nutrients in feeds: availability of major mineral ions. J. Anim. Sci. 35(3) p.695-712, 1972. PEO JR, E.R. Calcium, phosphorus and vitamin D in swine nutrition. IN: MILLER, E.R; ULLREY, D.E.; LEWIS, A.J. Swine nutrition. Butterworth-Neinemann, Stoneham, p.165182, 1991. ROSOL, T.J,; CHEW, D.J.; NAGODE, L.A.; CAPEN, C.C. Pathophysiology of calcium metabolism. Vet. Clin Ph. 24(2) p.49-63, 1995. SCOTT, T.A.; KAMPEN, R.; SILVERSIDES, F.G. The effect of phosphorus, phytase enzyme, and calcium on the performance of layers fed corn-based diets. Poult. Sci. 78, p.1742-1749, 1999. VRZUGULA, L. Metabolic disorders and their prevention in farm animals. Elsevier: Amsterdam, 1991. 390p. (Development in Animal and Veterinary Sciences, 24). WILSON, R.P.; ROBINSON, E.H.; GATLIN III, D.M.; POE, W.E. Dietary phosphorus requirement of channel catfish. J. Nutr. , 112, p. 1197-1202, 1982.
201
Anda mungkin juga menyukai
- 365594-Apostila de Nutriã Ã o Prof (1) - WalterDokumen201 halaman365594-Apostila de Nutriã Ã o Prof (1) - WalterEllen Ramos AmaralBelum ada peringkat
- Curso de Nutrição em Bovinocultura: Sistema DigestórioDokumen11 halamanCurso de Nutrição em Bovinocultura: Sistema DigestórioMoniqueMacedoBelum ada peringkat
- Digestão Geral Dos AnimaisDokumen17 halamanDigestão Geral Dos AnimaisThiago PortesBelum ada peringkat
- Anatomia Do Sistema DigestórioDokumen113 halamanAnatomia Do Sistema DigestórioMilena MarinhoBelum ada peringkat
- Obtenção de Matéria Pelos Seres HeterotróficosDokumen4 halamanObtenção de Matéria Pelos Seres HeterotróficosMariana DiasBelum ada peringkat
- Fisiologia Do Sistema Digestório em Animais Fisio 6Dokumen50 halamanFisiologia Do Sistema Digestório em Animais Fisio 6Maria Vitória Ferreira Ovídio100% (1)
- Avaliação - Nutrição de Não RuminantesDokumen4 halamanAvaliação - Nutrição de Não RuminantesERIKA APARECIDA SILVA NAVASBelum ada peringkat
- Anatomia Do Sistema DigestórioDokumen74 halamanAnatomia Do Sistema DigestórioBianca FranciscoBelum ada peringkat
- Fisiologia AnimalDokumen40 halamanFisiologia AnimalJosé Hilário Francisco da RosaBelum ada peringkat
- Sistema digestório: funções, processos e estruturaDokumen34 halamanSistema digestório: funções, processos e estruturaMaria Stella FernandesBelum ada peringkat
- Sistema Digestório - UnintaDokumen31 halamanSistema Digestório - UnintaEmily OliveiraBelum ada peringkat
- Sistema digestivo de aves, répteis e peixesDokumen30 halamanSistema digestivo de aves, répteis e peixesPriscila LessaBelum ada peringkat
- Sistema DigestórioDokumen24 halamanSistema DigestórioCamila BoechatBelum ada peringkat
- Sistema DigestórioDokumen7 halamanSistema DigestórioSaraiva JuniorBelum ada peringkat
- Resumo TgiDokumen4 halamanResumo TgiErycka Priscylla Gigante GarciaBelum ada peringkat
- Unidade 2Dokumen58 halamanUnidade 2Davi Barcelos SuttoffBelum ada peringkat
- Sistema Digestivo ResumoDokumen15 halamanSistema Digestivo ResumoTeótimo EduardoBelum ada peringkat
- Trato digestivo animaisDokumen66 halamanTrato digestivo animaisFabíola ArtemisBelum ada peringkat
- Anatomia - Módulo 3Dokumen26 halamanAnatomia - Módulo 3jhophaneBelum ada peringkat
- Fisiologia Do TGIDokumen34 halamanFisiologia Do TGILuciana SantosBelum ada peringkat
- Sistema digestório: órgãos, funções e processosDokumen9 halamanSistema digestório: órgãos, funções e processosMaria EduardaBelum ada peringkat
- Sistema Digestivo InsetosDokumen8 halamanSistema Digestivo InsetosTaynaBelum ada peringkat
- Sistema digestório: anatomia e funçõesDokumen81 halamanSistema digestório: anatomia e funçõesGeovanna Uchoa FurtadoBelum ada peringkat
- Digestão ExtracelularDokumen6 halamanDigestão ExtracelularFernando GalvãoBelum ada peringkat
- Resumos - Obtenção de Matéria Pelos Seres HeterotróficosDokumen7 halamanResumos - Obtenção de Matéria Pelos Seres HeterotróficosMatilde PedroBelum ada peringkat
- Aula 3Dokumen6 halamanAula 3Lisdalia CulandaBelum ada peringkat
- Resumo de Sistema DigestórioDokumen8 halamanResumo de Sistema DigestóriolaiskomatiBelum ada peringkat
- Fisiologia Da Digestão e Da Absorção em AvesDokumen6 halamanFisiologia Da Digestão e Da Absorção em AvesDaniela LimaBelum ada peringkat
- Fisiologia DigestivaDokumen26 halamanFisiologia DigestivaAlexandre Martins100% (1)
- Resumo Obtenção de Matéria e EnergiaDokumen11 halamanResumo Obtenção de Matéria e EnergiaSusana AlvesBelum ada peringkat
- 8º AnoDokumen15 halaman8º AnoDaniela SilvaBelum ada peringkat
- Sist. Digest. Das AvesDokumen4 halamanSist. Digest. Das Avessammyprado13Belum ada peringkat
- Nutrição BásicaDokumen4 halamanNutrição BásicaannielinutriBelum ada peringkat
- Fisiologia Dos InsetosDokumen16 halamanFisiologia Dos InsetosRaulBelum ada peringkat
- Funções Gerais Do Trato Gastrointestinal e Seu ControleDokumen2 halamanFunções Gerais Do Trato Gastrointestinal e Seu ControleNoé Da Sua LaiaBelum ada peringkat
- Fisiologia Do Sistema Digestório.P3Dokumen24 halamanFisiologia Do Sistema Digestório.P3thatsmarianaBelum ada peringkat
- TRABALHO ESCOLAR Sistema digestórioDokumen6 halamanTRABALHO ESCOLAR Sistema digestórioZeLuiz CruzBelum ada peringkat
- Processos digestivos em diferentes organismosDokumen9 halamanProcessos digestivos em diferentes organismosBernardo RepolhoBelum ada peringkat
- Resumo TGIDokumen5 halamanResumo TGIstock.anaaBelum ada peringkat
- ESTÔMAGODokumen20 halamanESTÔMAGOMichelle AraújoBelum ada peringkat
- Histologia E Embriologia: Adriana Dalpicolli RodriguesDokumen26 halamanHistologia E Embriologia: Adriana Dalpicolli Rodriguesjanainaaparecidadejesus22Belum ada peringkat
- aula 2 UNCDokumen20 halamanaula 2 UNCMsDebora ReinertBelum ada peringkat
- Anatomo Fisiologia Da DisgestãoDokumen7 halamanAnatomo Fisiologia Da DisgestãoBrunely OliveiraBelum ada peringkat
- FISIOLOGIA DA DIGESTÃO E ABSORÇÃO DE CARBOIDRATOS NAS AVES Renato - Furlan PDFDokumen18 halamanFISIOLOGIA DA DIGESTÃO E ABSORÇÃO DE CARBOIDRATOS NAS AVES Renato - Furlan PDFJuliana AlexandreBelum ada peringkat
- Estudo Dirigido - Problema 2 ASE 5Dokumen28 halamanEstudo Dirigido - Problema 2 ASE 5ANDRESSA DA CRUZ GURGELBelum ada peringkat
- Obtenção de matéria pelos seres vivos heterotróficosDokumen20 halamanObtenção de matéria pelos seres vivos heterotróficosLeticia VieiraBelum ada peringkat
- Nutrição, digestão e absorçãoDokumen4 halamanNutrição, digestão e absorçãoLauraBelum ada peringkat
- Aula 5. Bovinocultura - Nutrição de Ruminantes PDFDokumen76 halamanAula 5. Bovinocultura - Nutrição de Ruminantes PDFKely Vitória bezerra dos SantosBelum ada peringkat
- 8 - Particularidades Anatômicas Do Sistema Digestório de Animais Ruminantes e Não RuminantesDokumen5 halaman8 - Particularidades Anatômicas Do Sistema Digestório de Animais Ruminantes e Não RuminantesSílvia Cristina de AguiarBelum ada peringkat
- Digestão intra e extracelularDokumen23 halamanDigestão intra e extracelularSandra CardosoBelum ada peringkat
- Aula 8 - Anatomia - Sistema Digestório AlessandraDokumen27 halamanAula 8 - Anatomia - Sistema Digestório AlessandradonizetemartinsclaudemirBelum ada peringkat
- Fisiologia do Sistema DigestórioDokumen77 halamanFisiologia do Sistema DigestórioTata PetroffBelum ada peringkat
- Sistema digestório: funções, estruturas e processosDokumen24 halamanSistema digestório: funções, estruturas e processosKarla MoniqueBelum ada peringkat
- Fisiologia da digestão e absorção em avesDokumen6 halamanFisiologia da digestão e absorção em avesDaniela LimaBelum ada peringkat
- Sistema Digestivo 9º AnoDokumen5 halamanSistema Digestivo 9º AnoNoélia Paiva MorgadoBelum ada peringkat
- Cien 8Dokumen4 halamanCien 8eae giovanniBelum ada peringkat
- Digestão AvesDokumen39 halamanDigestão AvesPlínio RodriguesBelum ada peringkat
- Necessidades energéticas processo digestório!Dokumen3 halamanNecessidades energéticas processo digestório!Renata MotaBelum ada peringkat
- Aula 2 - Processo Digestivo AnimalDokumen62 halamanAula 2 - Processo Digestivo AnimalGislan Rocha100% (1)
- Ed 1 2013 DPF Administrativo AberturaDokumen32 halamanEd 1 2013 DPF Administrativo AberturaVinícius MeirelesBelum ada peringkat
- AREA 20 - 1 ParteDokumen6 halamanAREA 20 - 1 PartePaulo Henrique CavalcanteBelum ada peringkat
- SECRETARIA DE ESTADO DE ADMINISTRAÇÃO PÚBLICA DO DISTRITO FEDERAL Edital #01-SEAP - SEE, de 04 de Setembro de 2013Dokumen1 halamanSECRETARIA DE ESTADO DE ADMINISTRAÇÃO PÚBLICA DO DISTRITO FEDERAL Edital #01-SEAP - SEE, de 04 de Setembro de 2013Paulo Henrique CavalcanteBelum ada peringkat
- Mpu 2 13 Ed 5 2013 Mpu 2 13 Res Final Objetiva Prov Disc Analistaan Analista v2Dokumen60 halamanMpu 2 13 Ed 5 2013 Mpu 2 13 Res Final Objetiva Prov Disc Analistaan Analista v2Paulo Henrique CavalcanteBelum ada peringkat
- Livro ENAPDokumen346 halamanLivro ENAPwilsinhomadridBelum ada peringkat
- AREA 20 - 2 ParteDokumen6 halamanAREA 20 - 2 PartePaulo Henrique CavalcanteBelum ada peringkat
- Concurso público para Analista de Finanças e Controle da STNDokumen31 halamanConcurso público para Analista de Finanças e Controle da STNPaulo Henrique CavalcanteBelum ada peringkat
- Ações premiadas no 16o Concurso Inovação na Gestão PúblicaDokumen191 halamanAções premiadas no 16o Concurso Inovação na Gestão PúblicaPaulo Henrique CavalcanteBelum ada peringkat
- Memocards - Edital Mpu 2013Dokumen61 halamanMemocards - Edital Mpu 2013Memocards BrasilBelum ada peringkat
- Edital Concurso Senado 2008Dokumen28 halamanEdital Concurso Senado 2008Gil SantosBelum ada peringkat
- Edital - ESAFDokumen17 halamanEdital - ESAFFábio SeixasBelum ada peringkat
- Técnicas de Auditoria paraDokumen51 halamanTécnicas de Auditoria parasilvio100% (1)
- Normas orçamentárias e financeiras brasileirasDokumen139 halamanNormas orçamentárias e financeiras brasileirasrosalopesbarbosa50% (2)
- Aspectos JuridicosDokumen69 halamanAspectos JuridicosLimaPereiraBelum ada peringkat
- Edital AuditorDokumen7 halamanEdital AuditordspousoBelum ada peringkat
- Rel AnalDokumen20 halamanRel Analevandro_carlosBelum ada peringkat
- Lista 8 DaviDokumen2 halamanLista 8 DaviwesllaneBelum ada peringkat
- Exercicios Teia AlimentarDokumen4 halamanExercicios Teia AlimentarLarissa Potratz HartwigBelum ada peringkat
- Estratégias de suplementação de bovinos em pastoDokumen18 halamanEstratégias de suplementação de bovinos em pastoAline BernardoBelum ada peringkat
- Apostila Educação, Saúde e NutriçãoDokumen61 halamanApostila Educação, Saúde e NutriçãoIatiçara OliveiraBelum ada peringkat
- Aula prática de pesagem e medição de ingredientesDokumen2 halamanAula prática de pesagem e medição de ingredientesJéssica Teles100% (1)
- Receitas doces em potes e coposDokumen8 halamanReceitas doces em potes e coposAline AmorimBelum ada peringkat
- Raças bovinas brasileirasDokumen1 halamanRaças bovinas brasileirasSara Dornelles WeisBelum ada peringkat
- Alimentação saudável para idosos com diabetesDokumen30 halamanAlimentação saudável para idosos com diabetesMauro De BonisBelum ada peringkat
- Azia Gravida o Que Tomar - Pesquisa GoogleDokumen1 halamanAzia Gravida o Que Tomar - Pesquisa GoogleCaroline LeticiaBelum ada peringkat
- ScriptDokumen4 halamanScriptJokson AntonioBelum ada peringkat
- Controle de Produção - PadariaDokumen68 halamanControle de Produção - PadariaDangeloDGBelum ada peringkat
- A Cidade na LiteraturaDokumen16 halamanA Cidade na LiteraturaLuciano ViegasBelum ada peringkat
- Relatório de estágio no Hospital Cuf Infante SantoDokumen36 halamanRelatório de estágio no Hospital Cuf Infante SantoTainara MartinsBelum ada peringkat
- Receitas PDFDokumen276 halamanReceitas PDFNayara GiazzonBelum ada peringkat
- Aula 14 - Sotaques Do Brasil - Vocabulario RegionalDokumen4 halamanAula 14 - Sotaques Do Brasil - Vocabulario RegionalPROF DIEGO YOUVIPBelum ada peringkat
- 1 Aula - ElementosintroduzDokumen24 halaman1 Aula - ElementosintroduzsouzajossuBelum ada peringkat
- Simpatias para Curar AnimaisDokumen3 halamanSimpatias para Curar AnimaisOrgoneSkiesBelum ada peringkat
- Diálogos sobre moradia e vizinhançaDokumen17 halamanDiálogos sobre moradia e vizinhançaFabio DautonicoBelum ada peringkat
- Adolescentes bebem cada vez mais cedoDokumen6 halamanAdolescentes bebem cada vez mais cedoTiago PiresBelum ada peringkat
- Guia para emagrecer com corridaDokumen106 halamanGuia para emagrecer com corridaAlexandra Senna100% (1)
- Atividade 16 e 17 5° AnoDokumen11 halamanAtividade 16 e 17 5° Anomichelle lageBelum ada peringkat
- Cardápio de Bolos e Doces 2021-2Dokumen4 halamanCardápio de Bolos e Doces 2021-2Luís GustavoBelum ada peringkat
- Livro de Receitas Fiqfit 02Dokumen38 halamanLivro de Receitas Fiqfit 02GengyskanBelum ada peringkat
- Quimifol - Quimifol BoroDokumen3 halamanQuimifol - Quimifol BoroCutelaria ArtesanalBelum ada peringkat
- Inspeção Estabelecimentos GeladosDokumen23 halamanInspeção Estabelecimentos GeladosPreta LimaBelum ada peringkat
- Ervas - Dádiva Do Altíssimo (Guia Fitoterápico)Dokumen148 halamanErvas - Dádiva Do Altíssimo (Guia Fitoterápico)Jose helderBelum ada peringkat
- Prova de residência médica em pediatriaDokumen33 halamanProva de residência médica em pediatriaCamila LuzBelum ada peringkat
- As melhores massas, coberturas e recheiosDokumen74 halamanAs melhores massas, coberturas e recheiosLorena MaiaBelum ada peringkat
- Processos de purificaçãoDokumen111 halamanProcessos de purificaçãoVitor Jurciukonis CelescuekciBelum ada peringkat
- Relatório RefratometriaDokumen4 halamanRelatório RefratometriaVida JerushaBelum ada peringkat