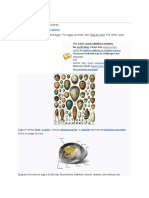
Anda mungkin juga menyukai
- Pigeons - A Collection of Articles on the Origins, Varieties and Methods of Pigeon KeepingDari EverandPigeons - A Collection of Articles on the Origins, Varieties and Methods of Pigeon KeepingBelum ada peringkat
- Anatomy ResearchDokumen8 halamanAnatomy Researchمحمد حامدBelum ada peringkat
- Sea Urchins as a Model for Studying Embryonic DevelopmentDokumen7 halamanSea Urchins as a Model for Studying Embryonic DevelopmentRed DiggerBelum ada peringkat
- Comparative Embryology, Aspects of Human Reproduction Pollution and Its Effect On OrganismsDokumen35 halamanComparative Embryology, Aspects of Human Reproduction Pollution and Its Effect On OrganismsSudesh RathodBelum ada peringkat
- 2011 Parental Care in FishesDokumen9 halaman2011 Parental Care in FishesHussnain ShahBelum ada peringkat
- DEVELOPMENTAL BIOLOGY 5: FERTILIZATION PROCESSDokumen12 halamanDEVELOPMENTAL BIOLOGY 5: FERTILIZATION PROCESSRajat K. SBelum ada peringkat
- Introduction To FertilizationDokumen4 halamanIntroduction To FertilizationSupermarc OlaguirBelum ada peringkat
- Structure of A Mature OvumDokumen6 halamanStructure of A Mature OvumZ/06 - Arko RoyBelum ada peringkat
- Reproduction: The Diversity of Fish Reproduction: An IntroductionDokumen3 halamanReproduction: The Diversity of Fish Reproduction: An IntroductioniqyuwidyaBelum ada peringkat
- Journal of Morphology - 2020 - Starck - Morphology of The Avian Yolk SacDokumen14 halamanJournal of Morphology - 2020 - Starck - Morphology of The Avian Yolk SacMashai LesenyehoBelum ada peringkat
- Bio 152 Lab 10 Animal Developemnt Worksheet PDFDokumen18 halamanBio 152 Lab 10 Animal Developemnt Worksheet PDFHyena100% (1)
- Marine Larval Fish Production A Nutritional Perspective.Dokumen12 halamanMarine Larval Fish Production A Nutritional Perspective.Hafez A MabroukBelum ada peringkat
- LAB EXERCISE 2 - OogenesisDokumen4 halamanLAB EXERCISE 2 - OogenesisGerald Angelo DeguinioBelum ada peringkat
- Class 8 Reproduction in Animals Notes of LessonDokumen11 halamanClass 8 Reproduction in Animals Notes of LessonSanthosh 456Belum ada peringkat
- Facultad de Ingeniería Marítima, Ciencias Biológicas, Oceánicas Y Recursos NaturalesDokumen5 halamanFacultad de Ingeniería Marítima, Ciencias Biológicas, Oceánicas Y Recursos NaturalesKleiner ArevaloBelum ada peringkat
- Animal Diversity and DevelopmentDokumen68 halamanAnimal Diversity and DevelopmentGabrielle Salamanca CastuloBelum ada peringkat
- Artificial Incubation of Fish Eggs: Proper Temperature, Water Flow are KeyDokumen5 halamanArtificial Incubation of Fish Eggs: Proper Temperature, Water Flow are KeyIvan H. OrtileBelum ada peringkat
- Vicilin-Derived Peptides Are Transferred From Males To Females As Seminal Nuptial Gift in The Seed-Feeding Beetle Callosobruchus MaculatusDokumen8 halamanVicilin-Derived Peptides Are Transferred From Males To Females As Seminal Nuptial Gift in The Seed-Feeding Beetle Callosobruchus MaculatusArianne FabresBelum ada peringkat
- Chick EmbryologyDokumen55 halamanChick EmbryologySushma MishraBelum ada peringkat
- Leche de CucarachaDokumen12 halamanLeche de CucarachaMARCO MARTIN SALAZAR BALDEONBelum ada peringkat
- PIIS1472648310616252Dokumen8 halamanPIIS1472648310616252PabloBelum ada peringkat
- The Sea Urchin Fertilization LabDokumen5 halamanThe Sea Urchin Fertilization LabAmy HollingsworthBelum ada peringkat
- Module 5 Heredity notes (1) HSC year 12Dokumen58 halamanModule 5 Heredity notes (1) HSC year 129yp4n7kxh8Belum ada peringkat
- Earth Science ModulesDokumen14 halamanEarth Science ModulesAlphaBelum ada peringkat
- Hemibagrus Nemurus: Ovarian Development of A River Catfish (Valenciennes, 1840) in CaptivityDokumen8 halamanHemibagrus Nemurus: Ovarian Development of A River Catfish (Valenciennes, 1840) in CaptivityCarlos YongBelum ada peringkat
- Early Ontogenesis of The Angelfish, Pterophyllum Scalare Schultze, 1823 (Cichlidae)Dokumen10 halamanEarly Ontogenesis of The Angelfish, Pterophyllum Scalare Schultze, 1823 (Cichlidae)kusuma wardanaBelum ada peringkat
- Bruce Miller-Early Life History of Marine Fishes-University of California Press (2009)Dokumen377 halamanBruce Miller-Early Life History of Marine Fishes-University of California Press (2009)Daniel Fernández100% (3)
- Fish reproduction organs and developmentDokumen11 halamanFish reproduction organs and developmentsksingl350Belum ada peringkat
- Class 11 Biology Zoology Em-Lesson 5 - Two Three Mark Study Material Englis PDFDokumen3 halamanClass 11 Biology Zoology Em-Lesson 5 - Two Three Mark Study Material Englis PDFuma1003Belum ada peringkat
- 1 Cephalopod Biology and Ongoing ResearchDokumen9 halaman1 Cephalopod Biology and Ongoing ResearchVicente VasquezBelum ada peringkat
- The Eggshell: Structure, Composition and Mineralization: Frontiers in Bioscience January 2012Dokumen16 halamanThe Eggshell: Structure, Composition and Mineralization: Frontiers in Bioscience January 2012John Carlo LayugBelum ada peringkat
- The Eggshell: Structure, Composition and Mineralization: Frontiers in Bioscience January 2012Dokumen16 halamanThe Eggshell: Structure, Composition and Mineralization: Frontiers in Bioscience January 2012Lagasca IrisBelum ada peringkat
- Els Lesson 2 Q2Dokumen24 halamanEls Lesson 2 Q2Febrian LarosaBelum ada peringkat
- M1 L2 wk2 HumanaDokumen19 halamanM1 L2 wk2 HumanaEdwin CaloloBelum ada peringkat
- Structures and Function of Fish Reproductive SystemDokumen8 halamanStructures and Function of Fish Reproductive Systemogunadeabiodun24Belum ada peringkat
- Penggunaan MikroskopDokumen6 halamanPenggunaan MikroskopKrisna Octa KBelum ada peringkat
- 8539Types of EggsDokumen8 halaman8539Types of Eggsouafia liiamaniBelum ada peringkat
- Blast UlaDokumen13 halamanBlast UlaOlif Ayu ZemiansyahBelum ada peringkat
- Principles of Development: Developmental BiologyDokumen86 halamanPrinciples of Development: Developmental BiologyPaolo OcampoBelum ada peringkat
- Organismos ModelosDokumen3 halamanOrganismos Modelosbrandy mirandaBelum ada peringkat
- Cells 10 02807 v2Dokumen25 halamanCells 10 02807 v2Wahyu PamungkasBelum ada peringkat
- Early Development of a South American FishDokumen9 halamanEarly Development of a South American FishDustin MitchellBelum ada peringkat
- In VitroDokumen10 halamanIn VitroJulian David Ortiz LondoñoBelum ada peringkat
- Comparative Vertebrate ReproDokumen6 halamanComparative Vertebrate ReproShaira CogollodoBelum ada peringkat
- Key Events in DevelopmentDokumen83 halamanKey Events in DevelopmentMelatiBelum ada peringkat
- Budding Is A Form of Asexual Reproduction That Results From The Outgrowth of ADokumen2 halamanBudding Is A Form of Asexual Reproduction That Results From The Outgrowth of AI Don't Know My NameBelum ada peringkat
- Role of Glycolysis and Respiration in Sperm Metabolism and MotilityDokumen60 halamanRole of Glycolysis and Respiration in Sperm Metabolism and MotilitywalczakcBelum ada peringkat
- Ratification Page: Suhardi Aldi Sri Hartini Nur ID. 1614042011 1414440008Dokumen16 halamanRatification Page: Suhardi Aldi Sri Hartini Nur ID. 1614042011 1414440008Hikmah Nur FadillahBelum ada peringkat
- Differences between external and internal fertilization in animalsDokumen2 halamanDifferences between external and internal fertilization in animalsLisly DizonBelum ada peringkat
- Artificial Incubation of Fish EggsDokumen5 halamanArtificial Incubation of Fish EggsClinton VillaflorBelum ada peringkat
- Jump To Navigationjump To Search: This Article Is About Biological Eggs. For Eggs As Food, See - For Other Uses, SeeDokumen5 halamanJump To Navigationjump To Search: This Article Is About Biological Eggs. For Eggs As Food, See - For Other Uses, SeeewbBelum ada peringkat
- Types of Eggs Submitted by T.Lakshanaa 3rd - Microbiology-"A"Dokumen3 halamanTypes of Eggs Submitted by T.Lakshanaa 3rd - Microbiology-"A"anjanaathilakar 05Belum ada peringkat
- Embryonic Development and Artificial Insemination of Clearnose SkatesDokumen17 halamanEmbryonic Development and Artificial Insemination of Clearnose SkatesJosé Vélez TacuriBelum ada peringkat
- SL Revised Viviparity DefinitionDokumen6 halamanSL Revised Viviparity Definitionapi-510500432Belum ada peringkat
- Special LessonDokumen73 halamanSpecial LessonJustine Mangrobang MendozaBelum ada peringkat
- Mini Review ArticleDokumen7 halamanMini Review ArticleEsabel LlanoBelum ada peringkat
- GASTRULATIONDokumen4 halamanGASTRULATIONProjjal SanyalBelum ada peringkat
- Study Mammalian Placenta TypesDokumen6 halamanStudy Mammalian Placenta TypesPrabhatBelum ada peringkat
- Zo 103Dokumen5 halamanZo 103Ravi ChauhanBelum ada peringkat
- Egg As Food Egg (Disambiguation) : Jump To Navigation Jump To SearchDokumen7 halamanEgg As Food Egg (Disambiguation) : Jump To Navigation Jump To Searchshreyaz 10Belum ada peringkat
- Lux Genes NSF Funding New Light and Life in Biology Teaching Laboratories Mini.4.CorsbyDokumen2 halamanLux Genes NSF Funding New Light and Life in Biology Teaching Laboratories Mini.4.CorsbyCHRISTEROPBelum ada peringkat
- Flathead Pursue Diversity Beyond The Phases of Meiosis VignaDokumen1 halamanFlathead Pursue Diversity Beyond The Phases of Meiosis VignaCHRISTEROPBelum ada peringkat
- Variation Inv Sexually Dimorphic Trait in The Human Skeleton V31reprintDokumen8 halamanVariation Inv Sexually Dimorphic Trait in The Human Skeleton V31reprintCHRISTEROPBelum ada peringkat
- An Introduction To Phylogenetic Analysis - Clemson UniversityDokumen51 halamanAn Introduction To Phylogenetic Analysis - Clemson UniversityprasadburangeBelum ada peringkat
- MillsDokumen2 halamanMillsMauricio MauricioBelum ada peringkat
- Lighting Up Chemiluminescent Southern Blot Procedure 2-CholewiakDokumen29 halamanLighting Up Chemiluminescent Southern Blot Procedure 2-CholewiakCHRISTEROPBelum ada peringkat
- Microscopy of Natural and Experimental Biofilms Simple Collection Kit 5-HenkDokumen18 halamanMicroscopy of Natural and Experimental Biofilms Simple Collection Kit 5-HenkCHRISTEROPBelum ada peringkat
- IslandBiogeography Students Colonize Islands To Test Hypotheses 12-HaefnerDokumen29 halamanIslandBiogeography Students Colonize Islands To Test Hypotheses 12-HaefnerCHRISTEROPBelum ada peringkat
- Microbial Ecology of The Oral Cavity 10-DillDokumen1 halamanMicrobial Ecology of The Oral Cavity 10-DillCHRISTEROPBelum ada peringkat
- Student-Directed Investigations Ofthe Actin Cytoskeleton Hamster Ovary V31reprintDokumen8 halamanStudent-Directed Investigations Ofthe Actin Cytoskeleton Hamster Ovary V31reprintCHRISTEROPBelum ada peringkat
- Stream Ecology and Water Quality Field Test Methods V31reprintDokumen17 halamanStream Ecology and Water Quality Field Test Methods V31reprintCHRISTEROPBelum ada peringkat
- Post It Note Science DNA Replication, Transcription and Translation Mini.1.Kreitzer-houslerDokumen4 halamanPost It Note Science DNA Replication, Transcription and Translation Mini.1.Kreitzer-houslerCHRISTEROP0% (1)
- Human Cardiopulmonary Laboratory 6-McfaddenDokumen16 halamanHuman Cardiopulmonary Laboratory 6-McfaddenCHRISTEROPBelum ada peringkat
- Offering Academic Assistance To Students in Your Biology Lab V31reprintDokumen9 halamanOffering Academic Assistance To Students in Your Biology Lab V31reprintCHRISTEROPBelum ada peringkat
- Using Reverse Genetics To Learn The Biological Function of A Gene V31reprintDokumen12 halamanUsing Reverse Genetics To Learn The Biological Function of A Gene V31reprintCHRISTEROPBelum ada peringkat
- Notes PDFDokumen18 halamanNotes PDFRohan EapenBelum ada peringkat
- Competition Within and Between Species of Parasitoid Wasps 12-GuinanDokumen21 halamanCompetition Within and Between Species of Parasitoid Wasps 12-GuinanCHRISTEROPBelum ada peringkat
- Purification of Maltose-Binding Protein From E. Coli Periplasm 3-LevinDokumen20 halamanPurification of Maltose-Binding Protein From E. Coli Periplasm 3-LevinCHRISTEROPBelum ada peringkat
- Biomechanical Analysis of Vertebrate Skeletal Systems 6-GlaseDokumen30 halamanBiomechanical Analysis of Vertebrate Skeletal Systems 6-GlaseCHRISTEROPBelum ada peringkat
- What's A Lab For Working Together To Optimize Lab Experience V31reprintDokumen32 halamanWhat's A Lab For Working Together To Optimize Lab Experience V31reprintCHRISTEROPBelum ada peringkat
- Teaching Western Blots With T Antigen and p53 02-GurneyDokumen24 halamanTeaching Western Blots With T Antigen and p53 02-GurneyCHRISTEROPBelum ada peringkat
- Gene Linkage & Mapping Using Tetrad Analysis in The Fungus Sordaria Fimicola 1-GlaseDokumen24 halamanGene Linkage & Mapping Using Tetrad Analysis in The Fungus Sordaria Fimicola 1-GlaseCHRISTEROPBelum ada peringkat
- Good The Bad Symbiotic Natural Populations of Wood Lice 7-NobletDokumen36 halamanGood The Bad Symbiotic Natural Populations of Wood Lice 7-NobletCHRISTEROPBelum ada peringkat
- 8 FogleDokumen22 halaman8 FoglePrashant Saxena100% (1)
- Photosynthetic Strategies Consequences For Community Succession 13-WinnettMurrayDokumen23 halamanPhotosynthetic Strategies Consequences For Community Succession 13-WinnettMurrayCHRISTEROPBelum ada peringkat
- Clearing Techniques Study of Vascular Plant Tissues 5-HerrDokumen23 halamanClearing Techniques Study of Vascular Plant Tissues 5-HerrCHRISTEROPBelum ada peringkat
- Using 3D Water Models To Investigate Fundamental Powerful Concepts Biology DemersDokumen8 halamanUsing 3D Water Models To Investigate Fundamental Powerful Concepts Biology DemersCHRISTEROPBelum ada peringkat
- 9 YeungDokumen19 halaman9 YeungNathan PittmanBelum ada peringkat
- Toolbox For Working With Living Vertebrates 19-DrewesDokumen15 halamanToolbox For Working With Living Vertebrates 19-DrewesCHRISTEROPBelum ada peringkat
- Developmental Biology of Frog Egg and SpermDokumen21 halamanDevelopmental Biology of Frog Egg and SpermRavindra MadurBelum ada peringkat
- Cleavage, Blastulation and Gastrulation Lab ReportDokumen8 halamanCleavage, Blastulation and Gastrulation Lab ReportJOSHUA MOLO100% (1)
- Cleavage & BlasulasiDokumen20 halamanCleavage & BlasulasiAkbar FadhlurrahmanBelum ada peringkat
- Vi Semester B. SC Zoology Practicals Study of Embryological Slides Frog BlastulaDokumen11 halamanVi Semester B. SC Zoology Practicals Study of Embryological Slides Frog BlastulaSuchitra SharmaBelum ada peringkat
- A Critical Appraisal of Garbha Avkranti Vis-A-VisDokumen10 halamanA Critical Appraisal of Garbha Avkranti Vis-A-VisAmitabh BachhanBelum ada peringkat
- Frog EmbryosDokumen43 halamanFrog EmbryosNics Martinez100% (7)
- ADokumen40 halamanAapi-3826751Belum ada peringkat
- B2603 ANIMAL DEVELOPMENT - Post Fertilization EventsDokumen13 halamanB2603 ANIMAL DEVELOPMENT - Post Fertilization EventssispulieBelum ada peringkat
- Development of starfish from fertilized egg to bipinarria larvaDokumen5 halamanDevelopment of starfish from fertilized egg to bipinarria larvaCamille100% (1)
- Embryogenesis 1Dokumen7 halamanEmbryogenesis 1api-319386998Belum ada peringkat
- Republic of The PhilippinesDokumen5 halamanRepublic of The Philippineshatdog adadaBelum ada peringkat
- Post Embryonic Development in BirdsDokumen11 halamanPost Embryonic Development in Birdshafsa yaqoobBelum ada peringkat
- Descriptive Embryology PDFDokumen12 halamanDescriptive Embryology PDFMuhammad KhalilBelum ada peringkat
- IETS Foundation Bovine in Vivo Ova Tutorial 2010 RevisionDokumen129 halamanIETS Foundation Bovine in Vivo Ova Tutorial 2010 RevisionHans Gallegos Rosado100% (2)
- Embryology Notes emDokumen25 halamanEmbryology Notes emAnonymous IwWT90VyBelum ada peringkat
- Lecture 16 Fate Mapping and Gastrulation in Sea UrchinDokumen23 halamanLecture 16 Fate Mapping and Gastrulation in Sea UrchinIQra KanwalBelum ada peringkat
- Early DevelopmentDokumen2 halamanEarly DevelopmentyelloweverglowBelum ada peringkat
- The Early Development of Chick: Cleavage & Gastrulation.: January 2015Dokumen33 halamanThe Early Development of Chick: Cleavage & Gastrulation.: January 2015dewiBelum ada peringkat
- Unit 13 Cleavage and GastrulationDokumen26 halamanUnit 13 Cleavage and GastrulationFERHAEEZA KALAYAKANBelum ada peringkat
- Exercise 3 - Embryology LabDokumen3 halamanExercise 3 - Embryology LabIvy CruzBelum ada peringkat
- 07 Gastrulation BiologyDokumen22 halaman07 Gastrulation BiologyKashif JamalBelum ada peringkat
- Practical I Section Iii: Embryology (1) Egg of InsectDokumen6 halamanPractical I Section Iii: Embryology (1) Egg of InsectAfridi ShaikhBelum ada peringkat
- The International Glossary On Infertility and Fertility Care, 2017Dokumen14 halamanThe International Glossary On Infertility and Fertility Care, 2017Zurya UdayanaBelum ada peringkat
- Embryogenesis 1Dokumen4 halamanEmbryogenesis 1kopipaym8Belum ada peringkat
- Essay Writers ServiceDokumen4 halamanEssay Writers Serviceafibafftauhxeh100% (2)
- Developmental BiologyDokumen43 halamanDevelopmental BiologyChala KelbessaBelum ada peringkat
- Developmental Biology Exam 1 Quiz 1 2 CH 13 Online Book Drosophila Online BookDokumen10 halamanDevelopmental Biology Exam 1 Quiz 1 2 CH 13 Online Book Drosophila Online BookPearl PascuaBelum ada peringkat
- Embryology Lecture Notes - 2 Cleavage and Types: The Planes of CleavageDokumen6 halamanEmbryology Lecture Notes - 2 Cleavage and Types: The Planes of CleavageQurat Ul EynBelum ada peringkat
- Embryology Study of Frog DevelopmentDokumen11 halamanEmbryology Study of Frog DevelopmentShree Mena OrnopiaBelum ada peringkat
- Embryology Study of Frog DevelopmentDokumen10 halamanEmbryology Study of Frog DevelopmentJerjer KingBelum ada peringkat
- Why We Die: The New Science of Aging and the Quest for ImmortalityDari EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityPenilaian: 3.5 dari 5 bintang3.5/5 (2)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDari EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisPenilaian: 3.5 dari 5 bintang3.5/5 (2)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDari EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RacePenilaian: 4.5 dari 5 bintang4.5/5 (515)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDari EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsPenilaian: 4.5 dari 5 bintang4.5/5 (4)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyDari EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyPenilaian: 3.5 dari 5 bintang3.5/5 (31)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorDari EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorBelum ada peringkat
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindDari EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindPenilaian: 4.5 dari 5 bintang4.5/5 (93)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondDari EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondPenilaian: 4 dari 5 bintang4/5 (3)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeDari EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeBelum ada peringkat
- The Lives of Bees: The Untold Story of the Honey Bee in the WildDari EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildPenilaian: 4.5 dari 5 bintang4.5/5 (44)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDari Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessPenilaian: 4 dari 5 bintang4/5 (33)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldDari EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldPenilaian: 4.5 dari 5 bintang4.5/5 (18)
- Why We Sleep: Unlocking the Power of Sleep and DreamsDari EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsPenilaian: 4.5 dari 5 bintang4.5/5 (2083)
- The Mind & The Brain: Neuroplasticity and the Power of Mental ForceDari EverandThe Mind & The Brain: Neuroplasticity and the Power of Mental ForceBelum ada peringkat
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesDari EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesPenilaian: 4.5 dari 5 bintang4.5/5 (396)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedDari EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedPenilaian: 4 dari 5 bintang4/5 (11)
- The Dragons of Eden: Speculations on the Evolution of Human IntelligenceDari EverandThe Dragons of Eden: Speculations on the Evolution of Human IntelligencePenilaian: 4 dari 5 bintang4/5 (632)
- Superlative: The Biology of ExtremesDari EverandSuperlative: The Biology of ExtremesPenilaian: 4.5 dari 5 bintang4.5/5 (51)
- Human Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesDari EverandHuman Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesPenilaian: 3.5 dari 5 bintang3.5/5 (56)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeDari EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifePenilaian: 4 dari 5 bintang4/5 (523)
- Summary of Robert M. Sapolsky's DeterminedDari EverandSummary of Robert M. Sapolsky's DeterminedPenilaian: 5 dari 5 bintang5/5 (2)
- Lymph & Longevity: The Untapped Secret to HealthDari EverandLymph & Longevity: The Untapped Secret to HealthPenilaian: 4.5 dari 5 bintang4.5/5 (13)
- Inside of a Dog: What Dogs See, Smell, and KnowDari EverandInside of a Dog: What Dogs See, Smell, and KnowPenilaian: 4 dari 5 bintang4/5 (390)
- Fearfully and Wonderfully: The Marvel of Bearing God's ImageDari EverandFearfully and Wonderfully: The Marvel of Bearing God's ImagePenilaian: 5 dari 5 bintang5/5 (40)
- Mitochondria and the Future of Medicine: The Key to Understanding Disease, Chronic Illness, Aging, and Life ItselfDari EverandMitochondria and the Future of Medicine: The Key to Understanding Disease, Chronic Illness, Aging, and Life ItselfPenilaian: 4.5 dari 5 bintang4.5/5 (98)
- Gathering Moss: A Natural and Cultural History of MossesDari EverandGathering Moss: A Natural and Cultural History of MossesPenilaian: 4.5 dari 5 bintang4.5/5 (347)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsDari EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsBelum ada peringkat
- Younger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationDari EverandYounger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationPenilaian: 4 dari 5 bintang4/5 (1)