Anda mungkin juga menyukai
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The Bible Does Not Condemn Premarital SexDokumen16 halamanThe Bible Does Not Condemn Premarital SexKeith502100% (3)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- Jesus Christ Was A HinduDokumen168 halamanJesus Christ Was A Hinduhbk22198783% (12)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- Internship Report On Effects of Promotion System On Employee Job Satisfaction of Janata Bank Ltd.Dokumen57 halamanInternship Report On Effects of Promotion System On Employee Job Satisfaction of Janata Bank Ltd.Tareq Alam100% (1)
- The Integumentary System Development: Biene, Ellen Angelic Flores, Andrie BonDokumen29 halamanThe Integumentary System Development: Biene, Ellen Angelic Flores, Andrie BonMu Lok100% (3)
- Corporation Law Case Digests Philippines Merger and ConsolidationDokumen7 halamanCorporation Law Case Digests Philippines Merger and ConsolidationAlpha BetaBelum ada peringkat
- Measures-English, Metric, and Equivalents PDFDokumen1 halamanMeasures-English, Metric, and Equivalents PDFluz adolfoBelum ada peringkat
- ThermodynamicsDokumen341 halamanThermodynamicsjonathan2788100% (4)
- 1990 Pekoz Design of Cold-Formed Steel Screw ConnectionsDokumen15 halaman1990 Pekoz Design of Cold-Formed Steel Screw ConnectionsmabuhamdBelum ada peringkat
- A Scenario of Cross-Cultural CommunicationDokumen6 halamanA Scenario of Cross-Cultural CommunicationN Karina HakmanBelum ada peringkat
- BSCHMCTT 101Dokumen308 halamanBSCHMCTT 101JITTUBelum ada peringkat
- Nguyễn Thị Ngọc Huyền - 19125516 - Homework 3Dokumen7 halamanNguyễn Thị Ngọc Huyền - 19125516 - Homework 3Nguyễn HuyềnBelum ada peringkat
- Contracts ChecklistDokumen3 halamanContracts ChecklistSteve WatmoreBelum ada peringkat
- Articles 62 & 63: Presented By: Muhammad Saad Umar FROM: BS (ACF) - B 2K20Dokumen10 halamanArticles 62 & 63: Presented By: Muhammad Saad Umar FROM: BS (ACF) - B 2K20Muhammad Saad UmarBelum ada peringkat
- Biometric Security ConcernsDokumen27 halamanBiometric Security ConcernsprinceuchenduBelum ada peringkat
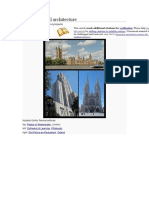
- Gothic Revival ArchitectureDokumen19 halamanGothic Revival ArchitectureAlexandra Maria NeaguBelum ada peringkat
- Argumentative Essay Project DescriptionDokumen5 halamanArgumentative Essay Project DescriptionKaren Jh MoncayoBelum ada peringkat
- The Role of Financial System in DevelopmentDokumen5 halamanThe Role of Financial System in DevelopmentCritical ThinkerBelum ada peringkat
- Second Grading EappDokumen2 halamanSecond Grading EappConnieRoseRamos100% (2)
- Defending A Dogma: Between Grice, Strawson and Quine: Elvis ImafidonDokumen10 halamanDefending A Dogma: Between Grice, Strawson and Quine: Elvis ImafidonYang Wen-LiBelum ada peringkat
- The ReformationDokumen20 halamanThe ReformationIlyes FerenczBelum ada peringkat
- Case Digest GR No 90501 NetDokumen4 halamanCase Digest GR No 90501 Netlawnotesnijan100% (1)
- Challenges Faced by Freight Forwarders in Their Operations in Chennai City, Tamil NaduDokumen3 halamanChallenges Faced by Freight Forwarders in Their Operations in Chennai City, Tamil NaduNiraj KasbeBelum ada peringkat
- Endocrine System Unit ExamDokumen3 halamanEndocrine System Unit ExamCHRISTINE JULIANEBelum ada peringkat
- Who, Summary NotesDokumen12 halamanWho, Summary NotesIvan Lohr100% (2)
- J of Cosmetic Dermatology - 2019 - Zhang - A Cream of Herbal Mixture To Improve MelasmaDokumen8 halamanJ of Cosmetic Dermatology - 2019 - Zhang - A Cream of Herbal Mixture To Improve Melasmaemily emiBelum ada peringkat
- LET Facilitating Learning EDITED3Dokumen12 halamanLET Facilitating Learning EDITED3Likhaan PerformingArts HomeStudio100% (5)
- Mathematics in The Primary Curriculum: Uncorrected Proof - For Lecturer Review OnlyDokumen12 halamanMathematics in The Primary Curriculum: Uncorrected Proof - For Lecturer Review OnlyYekeen Luqman LanreBelum ada peringkat
- Unsaturated Polyester Resins: Influence of The Styrene Concentration On The Miscibility and Mechanical PropertiesDokumen5 halamanUnsaturated Polyester Resins: Influence of The Styrene Concentration On The Miscibility and Mechanical PropertiesMamoon ShahidBelum ada peringkat
- Entrepreneur: Job/Career InvolvementDokumen5 halamanEntrepreneur: Job/Career InvolvementYlaissa GeronimoBelum ada peringkat
- Modelling of Short Duration Isopluvial Map For Raichur District KarnatakaDokumen4 halamanModelling of Short Duration Isopluvial Map For Raichur District KarnatakaMohammed Badiuddin ParvezBelum ada peringkat