Anda mungkin juga menyukai
- Carotid Doppler Study: Dr.I.Gurubharath MD PHD DR - Pooja MDDokumen116 halamanCarotid Doppler Study: Dr.I.Gurubharath MD PHD DR - Pooja MDJing CruzBelum ada peringkat
- Sports ScreeningDokumen8 halamanSports ScreeningJing CruzBelum ada peringkat
- Core 5.1 14122017FDokumen3 halamanCore 5.1 14122017FJing CruzBelum ada peringkat
- 2020 in Review: A Focus On Members, An Emphasis On ActionDokumen1 halaman2020 in Review: A Focus On Members, An Emphasis On ActionJing CruzBelum ada peringkat
- Phlebolymphology: Vol 28 - No. 1 - 2021 - P1-36 No. 103Dokumen39 halamanPhlebolymphology: Vol 28 - No. 1 - 2021 - P1-36 No. 103Jing CruzBelum ada peringkat
- Restrictive CardiomyopathyDokumen15 halamanRestrictive CardiomyopathyJing CruzBelum ada peringkat
- Alternate Pathway PrerequisitesDokumen1 halamanAlternate Pathway PrerequisitesJing CruzBelum ada peringkat
- Credentialing Process Sonography Canada: Is The Application Approved?Dokumen1 halamanCredentialing Process Sonography Canada: Is The Application Approved?Jing CruzBelum ada peringkat
- ARVCDokumen7 halamanARVCJing CruzBelum ada peringkat
- Focused Cardiac Ultrasound: Recommendations From The American Society of EchocardiographyDokumen15 halamanFocused Cardiac Ultrasound: Recommendations From The American Society of EchocardiographyPaola CastellanosBelum ada peringkat
- A Guideline Protocol For The Diagnosis and Assessment of Marfans Syndrome From The British Society of Echocardiography Education CommitteeDokumen3 halamanA Guideline Protocol For The Diagnosis and Assessment of Marfans Syndrome From The British Society of Echocardiography Education CommitteeJing CruzBelum ada peringkat
- Mitral Valve RepairDokumen5 halamanMitral Valve RepairJing CruzBelum ada peringkat
- AIM Guidelines ArterialUltrasoundDokumen20 halamanAIM Guidelines ArterialUltrasoundSmithery DiaBelum ada peringkat
- 5 Upper Extremity Arterial Duplex Evaluation Updated 2018 PDFDokumen8 halaman5 Upper Extremity Arterial Duplex Evaluation Updated 2018 PDFHasan Mohammad NayeemBelum ada peringkat
- Implementation Rural AllowancesDokumen15 halamanImplementation Rural AllowancesJing CruzBelum ada peringkat
- Practice Guidelines For Central Venous AccessDokumen6 halamanPractice Guidelines For Central Venous AccessJing CruzBelum ada peringkat
- Vascular Technology Content OutlineDokumen5 halamanVascular Technology Content OutlineJing CruzBelum ada peringkat
- Pre Op Hemo ProtocolDokumen2 halamanPre Op Hemo ProtocolJing CruzBelum ada peringkat
- Pre Op Hemo ProtocolDokumen2 halamanPre Op Hemo ProtocolJing CruzBelum ada peringkat
- Policies and Statements: Peripheral Arterial UltrasoundDokumen5 halamanPolicies and Statements: Peripheral Arterial UltrasoundJing CruzBelum ada peringkat
- Clinical Assessment of Ultrasound Equipment Form PDFDokumen1 halamanClinical Assessment of Ultrasound Equipment Form PDFJing CruzBelum ada peringkat
- Practice Guidelines For Central Venous Access PDFDokumen35 halamanPractice Guidelines For Central Venous Access PDFJulio AltamiranoBelum ada peringkat
- Ultrasound Technician Resume Example (Grace Medical Clinic) - Quitaque, TexasDokumen4 halamanUltrasound Technician Resume Example (Grace Medical Clinic) - Quitaque, TexasJing Cruz0% (1)
- G1 Clinical Application Specialist Resume Example (HITACHI HEALTHCARE AMERICAS) - Cary, IllinoisDokumen5 halamanG1 Clinical Application Specialist Resume Example (HITACHI HEALTHCARE AMERICAS) - Cary, IllinoisJing CruzBelum ada peringkat
- URS Edu HIVAIDS Window HopeDokumen19 halamanURS Edu HIVAIDS Window HopeJing CruzBelum ada peringkat
- Ge Point of Care - General Electric UltrasoundDokumen4 halamanGe Point of Care - General Electric UltrasoundJing CruzBelum ada peringkat
- Ultrasound Technician Resume Example (Saint Mary Hospital OB Clinic) - Wilmington, CaliforniaDokumen4 halamanUltrasound Technician Resume Example (Saint Mary Hospital OB Clinic) - Wilmington, CaliforniaJing CruzBelum ada peringkat
- HRQ Common Interview QuestionsDokumen2 halamanHRQ Common Interview QuestionsJing CruzBelum ada peringkat
- Tools For Evaluating HIV PDFDokumen59 halamanTools For Evaluating HIV PDFJing CruzBelum ada peringkat
- Urs Edu - Hivaids BotswaniaDokumen129 halamanUrs Edu - Hivaids BotswaniaJing CruzBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- The TMJ, Muscles, Nerves, and Ligaments With Voice Over - PPSXDokumen24 halamanThe TMJ, Muscles, Nerves, and Ligaments With Voice Over - PPSXJANINE PASIONBelum ada peringkat
- The Bone Book-An Orthopedic Pocket Manual (2020)Dokumen234 halamanThe Bone Book-An Orthopedic Pocket Manual (2020)Mohammad IbrahimBelum ada peringkat
- MuscolinoDokumen6 halamanMuscolinojustnewaroundBelum ada peringkat
- Master Kinesiotaping HandoutDokumen11 halamanMaster Kinesiotaping HandoutcosasdeangelBelum ada peringkat
- OinaDokumen19 halamanOinaPauline JaleaBelum ada peringkat
- Abductor Muscles and Their FunctionDokumen2 halamanAbductor Muscles and Their Functionstargazerlilies07Belum ada peringkat
- Sliding Filament Model of ContractionDokumen3 halamanSliding Filament Model of ContractionRATU ZULFI AMALIAHBelum ada peringkat
- Trigger Point TherapyDokumen45 halamanTrigger Point TherapyDina Umarella PihangBelum ada peringkat
- MCQ - MuscleDokumen10 halamanMCQ - Musclebiruk GBelum ada peringkat
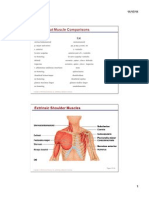
- Human/Cat Muscle Comparisons: Extrinsic Shoulder MusclesDokumen40 halamanHuman/Cat Muscle Comparisons: Extrinsic Shoulder MusclesJessica VanceBelum ada peringkat
- Hamstring MuscleDokumen8 halamanHamstring MuscleMuhamad SdeqBelum ada peringkat
- Stretching Exercise For Back and Lower BodyDokumen33 halamanStretching Exercise For Back and Lower BodyPhooi Yee Lau100% (3)
- Atlas of Human Anatomy - Including Student Consult Interactive Ancillaries and Guides1Dokumen1 halamanAtlas of Human Anatomy - Including Student Consult Interactive Ancillaries and Guides1JackTroviBelum ada peringkat
- Anatomy, Shoulder and Upper Limb, Shoulder MusclesDokumen12 halamanAnatomy, Shoulder and Upper Limb, Shoulder MusclesluizamgoBelum ada peringkat
- Síndrome Cruzado Superior e InferiorDokumen2 halamanSíndrome Cruzado Superior e InferiorFernandoBelum ada peringkat
- Anatomy Muscles of The FaceDokumen7 halamanAnatomy Muscles of The Facecristienne marielle MercadoBelum ada peringkat
- Skeletal Muscle StructureDokumen38 halamanSkeletal Muscle Structuretoyy100% (1)
- The Science: Self Myofascial Release-Foam RollingDokumen22 halamanThe Science: Self Myofascial Release-Foam RollingForminte MarianBelum ada peringkat
- Chapter 8 NotesDokumen4 halamanChapter 8 Notesbrady.anderson3017Belum ada peringkat
- Scapula - Elevation and DepressionDokumen16 halamanScapula - Elevation and DepressiondulexBelum ada peringkat
- Physio Muscle PhysiologyDokumen7 halamanPhysio Muscle PhysiologyApple AcaBelum ada peringkat
- Table Summary For Gross Anatomy of Upper LimbDokumen20 halamanTable Summary For Gross Anatomy of Upper Limbafifah zabidiBelum ada peringkat
- Muscular System Trans Week 1Dokumen9 halamanMuscular System Trans Week 1olaya.xyzanigaia.bsnBelum ada peringkat
- Anatomy of Fat Pads CotofanaDokumen15 halamanAnatomy of Fat Pads Cotofanadominique100% (2)
- Lower Limb Spotters: Hip & Gluteal Region, ThighDokumen6 halamanLower Limb Spotters: Hip & Gluteal Region, ThighDhaarshieth K R100% (1)
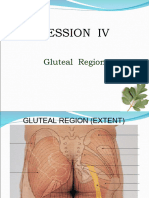
- Gluteal RegionDokumen27 halamanGluteal RegionRachita BhayekarBelum ada peringkat
- Engineering Approaches For Characterizing Soft Tissue Mechanical Properties - A Review PDFDokumen14 halamanEngineering Approaches For Characterizing Soft Tissue Mechanical Properties - A Review PDFdess101Belum ada peringkat
- Physioex Lala PDFDokumen5 halamanPhysioex Lala PDFNamira alifahBelum ada peringkat
- Table of Upper Limb MusclesDokumen7 halamanTable of Upper Limb MusclesLjubica Nikolic100% (7)
- The Course of N MedianusDokumen2 halamanThe Course of N MedianusResi Lystianto PutraBelum ada peringkat