Sem 3 Molecu
Diunggah oleh
StevenSilvaGodoy0 penilaian0% menganggap dokumen ini bermanfaat (0 suara)
33 tayangan12 halamanHak Cipta
© © All Rights Reserved
Format Tersedia
DOCX, PDF, TXT atau baca online dari Scribd
Bagikan dokumen Ini
Apakah menurut Anda dokumen ini bermanfaat?
Apakah konten ini tidak pantas?
Laporkan Dokumen IniHak Cipta:
© All Rights Reserved
Format Tersedia
Unduh sebagai DOCX, PDF, TXT atau baca online dari Scribd
0 penilaian0% menganggap dokumen ini bermanfaat (0 suara)
33 tayangan12 halamanSem 3 Molecu
Diunggah oleh
StevenSilvaGodoyHak Cipta:
© All Rights Reserved
Format Tersedia
Unduh sebagai DOCX, PDF, TXT atau baca online dari Scribd
Anda di halaman 1dari 12
Evolucin de operones trp bacteriana y su regulacin
Enrique Merino 1, Roy A Jensen 2, y Charles Yanofsky 3
1 Departamento de Microbiologa Molecular, Instituto de Biotecnologa de la
Universidad Nacional Autnoma de Mxico, Cuernavaca, Morelos 62271, Mxico
(merino@ibt.unam.mx)
2 Emerson Hall, PO Box 110700, Universidad de Florida, Gainesville, FL 32611,
EE.UU. (rjensen@ufl.edu)
Departamento de Ciencias Biolgicas, Universidad de Stanford, Stanford 3, CA 94305
a 5020, EE.UU. (Yanofsky@stanford.edu)
Resumen
La supervivencia y la replicacin de la mayora de las bacterias requieren la capacidad
de sintetizar el aminocido L-triptfano siempre que no est disponible desde el medio
ambiente. En este artculo se describen los genes, operones, protenas y reacciones
implicadas en la biosntesis de triptfano en las bacterias, y los mecanismos que
utilizan en la regulacin de la formacin de triptfano. Se demuestra que a pesar de
las reacciones de la biosntesis de triptfano son esencialmente idnticas, la
organizacin de genes vara entre las especies - de operones que toda-va a genes
dispersado completamente. Tambin muestran que los mecanismos de regulacin
utilizados para estos genes son muy variadas. Nos dirigimos a la pregunta - cules
son algunas de las ventajas potenciales de la variacin gen organizacin y la
regulacin asociada a esta conservada, va importante?
Introduccin
Existe un amplio conocimiento de los genes, las enzimas, operones y reacciones de L-
triptfano (Trp) biosntesis de las bacterias, y sobre los mecanismos que utilizan en la
regulacin de la formacin de Trp [** 1-5]. Todos los organismos con esta va utilizan
enzimas estructuralmente similares, lo que sugiere que los genes de esta va
evolucionaron slo una vez, probablemente tarde en la evolucin de los genes para la
biosntesis de aminocidos. Trp es uno de los aminocidos ms raras en la mayora de
las protenas, y es la ms costosa de sintetizar. Est codificado generalmente por un
solo codn, UGG, que puede haber servido como un codn de parada en la evolucin
codn temprano. Las estructuras 3D de los siete de las enzimas biosintticas de Trp
se conocen, y la mayora se asemejan a las estructuras de las protenas que catalizan
reacciones similares en otras vas. La organizacin de los genes trp dentro de
operones y los mecanismos de regulacin que se utilizan para controlar la expresin
del opern trp tanto varan mucho, sin duda, lo que refleja la divergencia del organismo
en relacin a los diferentes contextos metablicos de biosntesis de Trp. Varias
protenas trans y cis-actuando sitios y regiones-actuando se utilizan para regular la
expresin del opern trp. La variedad de mecanismos utilizados sugiere que los
grupos de organismos experimentaron diferentes presiones selectivas en respuesta a
las diferentes capacidades y necesidades. Las concentraciones intracelulares de Trp y
de cargados y no cargados ARNt Trp son las seales ms frecuentemente detectadas
en la regulacin del opern trp. En este artculo revisamos y actualizamos el
conocimiento sobre la distribucin de los genes, las enzimas y los mecanismos de
regulacin utilizados por las bacterias en la biosntesis de Trp.
Direccin del autor: Yanofsky, Charles (yanofsky@stanford.edu).
De la editorial Descargo de responsabilidad: Este es un archivo PDF de un manuscrito
indito que ha sido aceptado para su publicacin. Como un servicio a nuestros clientes
estamos proporcionando esta primera versin del manuscrito. El manuscrito ser
sometido a la correccin de estilo, composicin, y la revisin de la prueba resultante
antes de que se publique en su forma citable final. Tenga en cuenta que durante los
errores del proceso de produccin se pueden descubrir lo que podra afectar el
contenido, y todos los avisos legales que se aplican a la revista pertenecen.
Distribucin filogentico de organizacin gen trp y asociados factores de regulacin,
elementos, y mecanismos
La figura 1 muestra un rbol 16S rRNA de los organismos seleccionados que son un
cdigo de colores de acuerdo con los principales grupos filogenticos en las bacterias.
Aunque algunos estudios sobre la regulacin de genes trp en Archaea han aparecido
recientemente [6-9], las cuales exceden el alcance de esta revisin. Se muestra a la
derecha son la agrupacin -gen trp, trp fusiones -Gene y caractersticas
reglamentarias. Genes trp que se han especializado roles funcionales distintos de la
biosntesis de Trp primaria y que por lo tanto no estn regulados por Trp no se
muestran. Unos pocos patgenos (Haemophilus, Coxiella, y Propionibacterium) que
parecen estar en el proceso de prdida de la va Trp [4] son tambin se muestra. La
amplia variacin en los arreglos de genes dentro de operones trp y el repertorio de
mecanismos alternativos utilizados en la regulacin de la transcripcin de estos genes
(Fig. 2), ilustran la plasticidad que las bacterias deben haber tenido para la
optimizacin de la biosntesis de Trp.
Sensing Trp y ARNt Trp en bacterias Gram-negativas, Actinobacteridae y Deinococci
No es de extraar, en particular organizaciones de los genes y las estrategias de
regulacin estn a menudo presentes en las agrupaciones filogenticas relacionados.
Por ejemplo, en Escherichia coli y otra inferior-Gammaproteobacteria se organizan los
siete genes biosintticos trp dentro de una nica unidad transcripcional, el opern trp,
que siempre tiene una fusin de genes trpCF y que, en un grupo ms pequeo, tiene
una fusin trpGD (Fig. 1). En este grupo de bacterias de dos mecanismos principales
se utilizan para la regulacin de la transcripcin - una Trp deteccin y un segundo
sensor descargadas tRNA Trp. La iniciacin de la transcripcin del opern trp se
regula por la represin por la protena activada por Trp TrpR represor (Figs. 2a-2b)
[10,11].
Reglamento por TrpR en Psychrobacter y Coxiella ha sido predicho por el anlisis
bioinformtico, y la regulacin por TrpR se ha demostrado experimentalmente [12] en
los Chlamydiales filogenticamente distantes (que no son bacterias Gram-negativas).
Hasta el momento, trpR general parece estar autoregulated, y existe
predominantemente en una monocistronic opern (Fig. 1). Sin embargo, en algunos
organismos trpR es co-transcrito con otros genes trp, lo que permite la expresin
coordinada, por ejemplo, en el trpREG, trpRBA, y operones de trpRDCFBA
Cryohalolentis Psychrobacter, Chlamydia trachomatis y Chlamydia caviae,
respectivamente (figura 1).
Descargada ARNt Trp tambin es detectada por los mecanismos de atenuacin de la
transcripcin como una seal reguladora (Figs. 2c-2d) [11, * 13,14]. Curiosamente, a
pesar operones trp que contienen todos los genes trp estn comnmente sujetos a
regulacin por la atenuacin de la transcripcin, en algunos organismos este
mecanismo se utiliza para regular slo un subconjunto de los genes Trp (Fig. 1). Por lo
tanto, la atenuacin de la transcripcin se utiliza para regular los operones trpE y
trpGDC de Pseudomonas aeruginosa, el opern trpEG de algunas
Alphaproteobacteria (por ejemplo, Rhizobium etli) y Actinobacteridae (por ejemplo,
Streptomyces coelicolor), as como el opern trpE monocistronic de Deinococci (por
ejemplo,
Deinococcus radiodurans) (figura 1). En P. aeruginosa (P. y entomophila) la
transcripcin del opern trpBA es activada por la protena TRPI en respuesta a la
acumulacin de indoleglycerol fosfato (RICG), un biosinttica Trp intermedia [15,16]
(Figs. 2e-2f). RICG se acumula cada vez que la concentracin celular Trp es baja. Los
operones TRPI y trpBA de estos organismos se transcriben de manera divergente (Fig.
1) y su transcripcin est regulada por la unin de TRPI en sus regiones reguladoras
superpuestas (Figs. 2E-2F). Cuando la concentracin de Trp es alta, la concentracin
RICG es baja, y TRPI se une cerca de su promotor y autoregulates su propia sntesis.
Cuando la concentracin es baja y Trp RICG acumula, dos molculas de TRPI
activados se unen cerca del promotor trpBA, activando trpBA transcripcin (Figs. 2e-
2f) [15,16].
Bacterias Sensing Trp y ARNt Trp en Gram-positivas
Aunque las bacterias Gram-positivas tambin detectan tanto el nivel intracelular de
libre Trp y la disponibilidad de seales reguladoras como ARNt Trp cargadas, bastante
diferentes estrategias son utilizados por estos organismos en la regulacin del opern
biosinttico de trp (Fig. 1). Por ejemplo, en B. subtilis y algunos de sus parientes
cercanos, es necesario coordinar la expresin de los siete genes trp para la biosntesis
de Trp. Seis de estos genes estn agrupados en el opern trp, trpEDCFBA, que se
transcribe como parte de un supraoperon tambin contiene genes implicados en la
ruta aromtico comn y, en fenilalanina, tirosina, y la biosntesis de histidina. El
sptimo gen trp, trpg, (tambin funciona como PABA) se encuentra en el opern de
folato no enlazados. La transcripcin del opern trp de B. subtilis est regulado por la
atenuacin por el PRT-activado Trp unin al ARN protena de atenuacin, TRAP
[17,18] (Figs. 2g-2h). En presencia de exceso de Trp, predomina terminacin de la
transcripcin TRAP mediada, pero esas pocas transcripciones que se escapan de
terminacin estn sujetos a la regulacin de traduccin a travs de la formacin de
una estructura secundaria que bloquea el acceso ribosoma al sitio de unin al
ribosoma-trpE [19]. Expresin de trpg est regulada coordinadamente con los otros
genes trp por TRAP activado-Trp, que se une en el sitio de unin de ribosomas trpg
ARNm, inhibiendo su traduccin [20]. Regulacin TRAP del opern trp tambin ocurre
en algunos clostridios, aunque en este caso la organizacin opern trp vara
significativamente de la de B. subtilis. Por ejemplo, en Syntrophomonas wolfei y
Carboxydothermus hydrogenoformans todos los genes trp se transcriben como parte
de un comn opern. En el primero, trpE y trpg se fusionan y situados en el extremo
del opern mientras que en este ltimo, el opern comienza con el gen aroF, seguido
por trpE no fusionado y genes trpg (Fig. 1).
En B. subtilis la disponibilidad de descargadas tRNA Trp tambin se detecta, y la
acumulacin de descargadas tRNA Trp regula la sntesis de AT, una protena anti-
TRAP. AT se une a TRAP activado-Trp, inhibiendo su funcin [21,22]. (Figs. 2i-2j). El
gen estructural para AT, RTPA, est regulado transcripcionalmente por el mecanismo
de caja de T (vase ms adelante), en respuesta a la acumulacin de tRNA no
cargados Trp [22]. AT sntesis tambin est regulada por traslacin descargadas tRNA
Trp [21]. AT parece haber evolucionado muy poco, ya que slo se ha identificado en B.
B. subtilis y el estrechamente relacionado licheniformis (Fig. 1). En los otros
organismos que contienen TRAP no se sabe si algn otro mecanismo de regulacin se
utiliza para detectar la acumulacin de tRNA no cargados Trp.
En la gran mayora de las bacterias Gram-positivas, TRAP no est presente. La
transcripcin del opern trp se regula en respuesta a la acumulacin de ARNt Trp sin
carga, por el mecanismo de caja T [23] (Figs. 2k-2l). Este mecanismo implica el
reconocimiento de ARN lder descargadas tRNA Trp - dirigido con gran especificidad y
afinidad en la ausencia de factores proteicos. El cuadro de T se identific
originalmente como el elemento regulador utilizado durante muchos operones
aminoacil-tRNA sintetasa en bacterias Gram-positivas [23]. Ms tarde se encontr que
se asocia con muchos otros operones del metabolismo de aminocidos, tales como los
que contienen genes transportadores y genes de la biosntesis. Los elementos de caja
T que regulan trp genes biosintticos son a menudo dispuestas en tndem en la regin
aguas arriba transcrita del opern trp. Sin embargo, en algunas especies slo un nico
elemento de caja T est presente [1 **, ** 24] Fig. 1). En los organismos que utilizan el
mecanismo de caja de T para detectar descargadas tRNA Trp en la regulacin de la
expresin del opern trp, la existencia de un mecanismo de deteccin de libre Trp es
desconocido. En los organismos con elementos de caja en tndem T, las ventajas
cuantitativos derivados de tndem empleando T cajas podran compensar la falta de
un mecanismo de regulacin por separado para detectar Trp [1 **, ** 24]. Reglamento
del opern trp por el mecanismo de la caja T se limita casi exclusivamente a los
Firmicutes.
Las posibles razones de las diferencias en la organizacin del opern y
mecanismos de regulacin
La va Trp requiere los productos de otras cuatro vas para la finalizacin de la
biosntesis de Trp. Corismato es el precursor comn de todos los tres aminocidos
aromticos, as como un precursor de otros metabolitos esenciales, por ejemplo, cido
flico. La glutamina proporciona un grupo amino esencial durante la formacin de
antranilato, PRPP es la fuente de varios tomos de carbono del anillo de indol de Trp,
y L-serina proporciona la cadena lateral de Trp. PRPP, un importante metabolito de
alta energa, debe ser utilizado de manera eficiente. Si antranilato exgeno fuera a
entrar en la clula y generar innecesaria Trp, esto podra disminuir la disponibilidad de
PRPP para la sntesis de L-histidina (y para otras funciones). . Esto probablemente
explica por qu transferasa fosforribosil antranilato es un segundo objetivo, adems de
la sintasa de antranilato (AS) para la inhibicin por retroalimentacin Trp en bacterias
entricas [25] Bacillus subtilis emplea una alternativa - e indirecta - mecanismo por el
cual su AS es activado por histidina [26] .
Ms all de la suerte de Trp como un sustrato para la sntesis de protenas, algunos
intermedios en la biosntesis de Trp - y el propio Trp - pueden servir otras funciones, y
estas funciones son altamente individualistas en la naturaleza. En vista de esto, tal vez
no sea sorprendente que la organizacin opern trp vara considerablemente en
diferentes especies bacterianas, y que los mecanismos diferentes se utilizan en la
regulacin de su expresin. (I) Los organismos como P. aeruginosa antranilato utilizar
para varios propsitos. (Vase la discusin posterior de la biosntesis de las
quinolonas en la seccin LGT). Por lo tanto, la necesidad global de AS aqu no puede
ser coordinado con la necesidad de la sntesis de otras enzimas de la ruta de Trp.
Estas relaciones pueden explicar en parte el opern split (Fig. 1) [4] y la regulacin
diferencial de las enzimas Trp visto en esta especie y sus parientes cercanos. (Ii) En
los organismos asociados de las plantas tales como Azospirillum brasilense [27] Trp
asiste planta crecimiento, al servir como precursor del cido indol-3-actico. Este
organismo posee dos como enzimas, uno sensible a la inhibicin por retroalimentacin
por Trp y controlado por una regin de pptido lder TrpL. El segundo AS no es ni
sensible a la inhibicin por retroalimentacin por Trp, ni se sabe que es objeto de
represin por Trp, sugerente de su funcin especializada en la sntesis de auxina [28].
(Iii) Chromobacterium violaceum produce grandes cantidades de pigmento, violacena,
que es derivado de dos molculas de Trp [29]. Los genes trp de este organismo (no
mostrados en la fig. 1) son en su mayora dispersos y presentes como copias nicas.
Sera interesante saber qu estrategias de control dictan los dos destinos de Trp en C.
violaceum. (iv) Streptomyces coelicolor posee un gran grupo de genes asociados con
la formacin de un antibitico dependiente de calcio que contiene Trp. El grupo incluye
copias de cuatro de los genes Trp (no mostrado en la fig. 1), lo que implica su
participacin especializada en la va de antibitico (4). Por lo tanto, los tres genes trp
restantes que estn presentes como copias nicas se utilizan nicamente para dos
funciones diferentes de la va. (V) especies de Cytophaga, Polaribacter, Xanthomonas,
y Gemmata (no mostrado en la fig. 1) sintetizan quinolinato de Trp y la utilizan como
un precursor de la niacina [30]. En algunos de estos organismos slo los genes trp
individuales estn presentes, lo que implica que existen mecanismos reguladores que
no dependen de la regulacin diferencial de mltiples copias de genes trp.
Un interesante alteracin de contexto para la va metablica Trp se ejemplifica por el
opern trp mosaico de Chlamydophila caviae (Fig. 1), un patgeno intracelular que
recicla quinurenina derivado de Trp del husped mamfero de nuevo a patgenos
disponible Trp [2]. El opern trp de este organismo carece de los genes que codifican
AS, pero posee los genes trp -pathway restantes. Su opern trp mosaico tambin
contiene genes que codifican PRPP sintetasa (no de otro modo presentes en la
Chlamydia) y kynureninase (necesario para la conversin de kynurinine anfitrin de
antranilato patgeno). El gen represor, trpR, est tambin dentro del opern, que
indica que es auto-regulado (Fig. 1).
Diversidad enzimtica
Las reacciones de la biosntesis de Trp son invariantes, pero las enzimas que catalizan
ellos a menudo presentan caractersticas distintas. Existen muchas combinaciones de
genes de fusin diferentes en diferentes linajes [4], y tales fusiones son marcadores
fiables de la relacin filogentica.
Antranilato sintasa (AS)
El polipptido TrpE, en complejo con el polipptido trpg, cataliza la formacin de
antranilato de corismato y glutamina [31]. P aminobenzoato (PABA) sintasa, una
estrecha homlogo de AS, lleva a cabo una reaccin similar para generar PABA para
la sntesis de folato [32]. Ambas enzimas utilizan una subunidad amidotransferasa
para entregar el amonaco derivado de glutamina al sitio activo. Como, pero no PABA
sintasa, es inhibida por retroalimentacin Trp. Desde PABA sintasa, sin embargo, tiene
un bolsillo Trp-unin, esto es consistente con un escenario evolutivo por el cual el gen
de la sintasa de PABA evolucion a partir de trpE por la duplicacin de genes, seguido
por la prdida de la inhibicin por retroalimentacin Trp. PABA no se deriva de
corismato en algunos organismos [* 33].
Fosforribosil transferasa antranilato
TrpD es una enzima altamente conservada, que junto con nuclesido fosforilasa y la
timidina fosforilasa representan uno de los tres tipos de pliegues fosfotransferasa [34].
Curiosamente, bajo ciertas condiciones, TrpD suministra amina fosforribosil para la
biosntesis de tiamina [35].
Los polipptidos TrpF, trpC, y TrpA
Estos tres enzimas son de tamao similar y todos han estructuras barril. Los tres
pueden haberse originado a travs de la duplicacin de genes a partir de un gen
ancestral que codifica una enzima que puede haber sido capaz de catalizar las tres
reacciones. En algunos actinomicetos la TrpF codificados con polipptido isomerasa
falta, y su funcin es proporcionada por el isomerasa similares histidina-va, denota
PriA [36]. El grupo de organismos que carecen de TrpF incluye toda la subclase
Actinobacteridae, excepto un pequeo grupo dentro de la Corynebacteriaceae Familia
(Fig. 1) en la que todo el opern trp (excepto por un remanente trpC) ha sido sustituido
por medio de la LGT (ver ms adelante).
TrpB
Algunos organismos tienen dos protenas trpB, uno de los cuales complejos con TrpA
y funciones en la formacin de Trp, mientras que el otro (a subhomolog claramente
distinto) puede actuar como una serina desaminasa [* 37,38]. Se ha propuesto que
TrpB y la enzima que cataliza el ltimo paso en la biosntesis de treonina tienen un
origen comn [39].
Transferencia lateral de genes
La transferencia de todo el camino y operones trp-va parcial a otros organismos a
travs de LGT ha sido ampliamente revisado recientemente [3,40]. Incluso cuando
LGT origen de un opern dado es claramente evidente, es raro que tanto el donante y
el receptor se han estudiado experimentalmente con el fin de deducir la evolucin de
roles funcionales alterados y regulacin alterada. Es interesante que las publicaciones
recientes permiten una comprensin global de opern trp LGT en los dos ejemplos
siguientes.
Whole-va LGT en Corynebacterium
Un ancestro comn de algunas de las especies dentro del gnero Corynebacterium
recibi un opern trp completa a partir de un miembro del linaje entrica [3,4]. Los
genes que tienen el papel funcional de la biosntesis de primaria Trp en el receptor han
sido reemplazados por genes que tienen el mismo papel funcional en el donante. El
genoma receptor se puede establecer claramente a un antepasado que exista hace
muy poco a la vez despus de la divergencia de C. Jeikeium (que no tiene el opern
LGT;. Fig 1), pero antes de la divergencia de C. glutamicum, C. efficiens, y C.
diptheriae (que todos tienen el opern LGT). Es muy interesante que la adquisicin del
opern LGT ha proporcionado la isomerasa codificada por TrpF, un gen que est de
otro modo ausente de un gran grupo de bacterias actinomicetos, como se discuti
previamente. Por lo tanto, como la mayora de los actinomicetos, C. jeikeium se basa
en la isomerasa histidina-va (PriA) para la doble funcin en dos vas, mientras que las
tres especies hermanas poseen isomerasas especializados (uno para la va de Trp y
uno para la va de histidina). El opern transferido tiene la fusin trpC / TrpF que es
tpico del linaje entrico. Elementos cis de regulacin que fueron transferidos incluyen
un pptido lder asociada con una regin atenuador, as como un promotor interno. El
gen estructural para el represor trpR no se transfiri, como podra esperarse de un gen
regulador disociados de los genes estructurales biosintticos. Sin embargo, una
publicacin reciente muestra que esto ha sido compensado por el servicio militar
obligatorio de LTBR, un represor transcripcional de tipo IclR [** 41] que tiene un papel
muy conservadas en la regulacin de la biosntesis de leucina en bacterias
actinomicetos. Por lo tanto, la va de Trp C. glutamicum puede ser apreciada como una
novela salto evolutivo que result de una combinacin de LGT de un donante distante
y reclutamiento de un gen regulador nativo.
LGT parcial-va en Pseudomonas aeruginosa
P. aeruginosa es una muy reciente destinatario de un opern trpEG de un donante
dentro del entrica linaje, es decir, el fragmento proximal del opern trp de siete gen.
Como se muestra por una publicacin reciente [** 42], esto es un ejemplo de un caso
en el que los genes, que en el donante se dedicaban a la biosntesis de primaria,
fueron reclutados para una nueva funcin especializada en el receptor (y por lo tanto
no se muestran en la fig. 1 donde slo los genes trp que participan en la biosntesis de
triptfano primaria se ilustran). Los elementos de control por delante de AS, o bien no
se transfieren o se desecharon. La sensibilidad a la inhibicin por retroalimentacin por
Trp tambin se perdi. El opern trpEG se coloc adyacente al opern pqsABCDE. En
conjunto, estos siete genes codifican los productos gnicos que producen 2-heptil-3-
hidroxi-4-quinolona (PQS), una percepcin de qurum de sealizacin compuesto y el
factor de virulencia. Los dos operones son controlados positivamente por PqsR, y los
productos de los genes se expresan principalmente durante el metabolismo fase
estacionaria. Por lo tanto, los genes trp aliengenas fueron asimilados en un contexto
funcional diferente y colocados bajo un rgimen de control completamente diferente.
Esto representa una innovacin muy reciente a nivel de especie ya que el opern LGT
no est presente en muy cercanos a P. aeruginosa. El evento LGT es suficientemente
reciente que se podra deducir que la especie donante trpEG quiz podran haber sido
de Vibrio.
Conclusiones
Los genes y operones de la ruta biosinttica de Trp se organizan de forma diferente en
diversas especies bacterianas. Estas diferencias reflejan divergencia evolutiva as
como el ajuste a las capacidades metablicas nicas y de las interacciones
medioambientales. Asociado con estas diferencias en la organizacin del opern son
el uso de una variedad de sitios que actan en cis y trans-actuando protenas en la
regulacin de la transcripcin del opern. Adems, algunos organismos pueden haber
adquirido, en un solo paso, a travs de LGT, un opern que ha experimentado
modificaciones evolutivas que le permiten ofrecer una nueva funcin y / o una nueva
regulacin. As LGT debe tambin haber contribuido a la diversidad opern que ahora
observamos.
Figura 1. Operon organizacin y regulacin de la transcripcin trp genes
trp opern organizacin y asociados factores de regulacin y elementos, estn
"pintadas" en un rbol filogentico de los organismos. Slo el Partido Radical
Transnacional genes que funcionan para la biosntesis de triptfano primaria, no PRT
genes que participan en las vas secundarias especializadas, se muestran. Las ramas
de Helicobacter pylori y Corynebacterium glutamicum estn codificados por colores
para indicar el origen LGT de sus todo-va el Partido Radical Transnacional operones.
Un alineamiento mltiple de las secuencias de 16S rRNA de genomas representativos
se utiliz para calcular las "distancias de similitud gentica" mediante el programa
PROTDIST del programa de paquete de inferencia filogenia PHYLIP. Sobre la base de
estas distancias estimadas, se realiz sucesivas agrupacin de linajes utilizando el
algoritmo de vecino a participar tal como se aplica en el programa VECINO. El ADN /
ARN regulador itios de unin, atenuadores de transcripcin, y pptidos lder indicados
se han identificado experimentalmente o predicho a partir de anlisis por ordenador de
las secuencias del genoma. Para grandes operones con ms de 5 no consecutivos
PRT genes (flechas blancas), se indica el nmero de estos genes. Los organismos con
un ARNt Trp -regulated T elemento de caja de control de la transcripcin de su trp
opern generalmente tambin contienen un ARNt Trp -regulated T elemento de caja
de control de la transcripcin de trpS , el gen estructural para triptofanoil-tRNA
sintetasa. mtrB, RTPA , y LTBR , son, respectivamente, los genes estructurales para la
trampa, polipptidos AT, y LtrB.
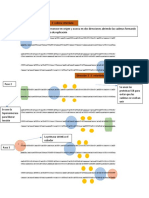
Figura 2. mecanismos moleculares utilizados en la regulacin de la transcripcin de los
genes de la trp opern biosinttico en bacterias
La variedad de factores y elementos que participan en la regulacin de la transcripcin
de trp operones biosintticos se muestran. Algunos bloquean la accin de la ARN
polimerasa, mientras que otros influyen en la terminacin prematura de la
transcripcin. Las seales que ms a menudo detectados son la concentracin
intracelular de libre Trp, y el nivel de descargadas tRNA Trp . Estos activan o regulan
la formacin de los diferentes componentes moleculares, incluyendo ADN y las
protenas de unin de ARN, ARN, protenas de unin a protenas, y los ribosomas de
traduccin. En algunos casos en los que la regulacin por terminacin de la
transcripcin se ha predicho, la seal asociada no se ha identificado. Regulacin de la
transcripcin basada en la activacin Trp del represor TrpR (a,b). La protena TrpR se
encuentra principalmente en Gammaproteobacteria y en algunas Chlamydiales (Fig.
1). (a) Cuando los niveles intracelulares de Trp son bajos, TrpR existe en su
conformacin inactiva aporrepresor que no puede unirse a trp ADN operador. En
consecuencia, la RNA polimerasa puede iniciar trp opern transcripcin. (b) Como el
nivel intracelular de Trp se eleva, se une a la TrpR aporrepresor, alterando su
conformacin. El represor se une en sus cognados trp operadores, la superposicin de
la trp promotor, la prevencin de la iniciacin de trp opern transcripcin [10].
atenuacin mediada por la transcripcin del ribosoma (c, d). En algunos organismos, el
nivel intracelular de ARNt cargado Trp determina si un ribosomas traducir se detendr
en uno de los dos adyacentes trpL codones Trp al intentar la sntesis del pptido lder
TrpL. (c) Bajo crecimiento condiciones donde el ARNt cargado Trp nivel es bajo, los
ribosomas traducir puestos en uno de los trpL codones de Trp. Ribosome
estancamiento favorece la formacin de la estructura antiterminator, en lugar de la
estructura terminador, por lo tanto, la transcripcin contina en el opern. (d) Cuando
el nivel de ARNt cargado Trp es alta, la traduccin de trpL se completa ARNm y el
ribosoma de traduccin se disocia. Esto permite la formacin de un terminador de la
transcripcin independiente de Rho en la transcripcin que finaliza la transcripcin en
la regin lder del opern. Reglamento basado en la activacin de la protena TRPI (e,
f) . La protena TRPI pertenece a la LysR-familia de protenas reguladoras procariotas.
Su gen estructural, TRPI , es adyacente a y transcritas de forma divergente desde el
trpBA opern que codifica las subunidades de los complejos enzima triptfano
sintetasa. (e) Cuando el nivel intracelular de Trp es baja, RICG, un intermedio
biosinttico y un sustrato de la sintasa de triptfano, se produce en exceso,
favoreciendo doble unin de TRPI al operador de los sitios en la TRPI-trpBA regin
intergnica. TRPI de unin activa la transcripcin de la trpBA opern, ya sea por
interaccin directa con la RNA polimerasa o mediante la induccin de ADN de flexin
[43].
(F) Bajo condiciones de crecimiento en donde existe un exceso de triptfano, la
concentracin RICG no es suficiente para inducir el reconocimiento de TRPI para su
sitio de unin de baja afinidad, por lo tanto la transcripcin de trpBA no est activado.
Reglamento de trp expresin gnica por TRPI parece estar restringida a
Pseudomonas aeruginosa , P. putida y P. syringae (Fig. 1). Reglamento sobre la base
de la accin de la protena de TRAP (g, h) . TRAP es una protena de unin al ARN se
encuentra en algunos Bacillales, incluyendo Bacillus subtilis , y en algunos clostridios
(Fig. 1). (g) Siempre que Trp es limitante del crecimiento, TRAP est inactivo. Cuando
TRAP no puede unirse a ARN, el ARN lder de la trp opern se pliega para formar la
estructura secundaria energticamente favorecida, la antiterminator. Esta estructura
impide la formacin de un terminador de la transcripcin independiente de Rho, en
consecuencia transcripcin procede en genes estructurales del opern. (h) Cuando hay un
exceso de triptfano, TRAP se activa, su preparacin para unirse al segmento de ARN lder de
la trp transcripcin del opern . Esta unin altera la estructura antiterminator, favoreciendo la
formacin de la estructura terminador, que conduce a la terminacin prematura de la
transcripcin en la regin lder del opern [17].
Reglamento sobre la base de la accin de la protena anti-TRAP AT (i, j). AT se
produce en respuesta a la acumulacin de tRNA no cargados. (i) Siempre que el
descargadas tRNA Trp es elevado nivel, AT se sintetiza y se une a Trp TRAP -
activated, inhibir la actividad de TRAP [44]. (j) Cuando el ARNt cargado Trp nivel es
alto, AT sntesis se evita, eliminando su efecto sobre la funcin TRAP [21,22].
Reglamento sobre la base de un elemento de caja T (k, l). (K) Cuando el ARNt
cargado Trp nivel es bajo, descargadas tRNA Trp estabiliza la estructura antiterminator
cuadro de T, prevencin de la formacin de la estructura terminador. Esto permite la
transcripcin para continuar en el resto del opern. (l) Cuando el ARNt cargado Trp
nivel es alto, este ARNt cargado es incapaz de estabilizar la estructura antiterminator.
El terminador, por lo tanto, las formas de terminacin de la transcripcin [23].
Anda mungkin juga menyukai
- Genómica en La Producción AnimalDokumen22 halamanGenómica en La Producción AnimalOmar Parrao OrdazBelum ada peringkat
- Adn y Replicación, Transcripción y Traducción.Dokumen32 halamanAdn y Replicación, Transcripción y Traducción.Paula SánchezBelum ada peringkat
- 10, No Menos ?Dokumen27 halaman10, No Menos ?angeljesusgarciafranco988Belum ada peringkat
- Biologìa Celular Resumen CBC Universidad de Buenos AiresDokumen39 halamanBiologìa Celular Resumen CBC Universidad de Buenos AiresKariu SolBelum ada peringkat
- El Flujo de La Información Genética. Guía 4Dokumen4 halamanEl Flujo de La Información Genética. Guía 4ApolinaVunoimaBelum ada peringkat
- Construcción de PlásmidosDokumen25 halamanConstrucción de PlásmidosJennifer Guevara FreireBelum ada peringkat
- Rejilla Capitulo 8Dokumen6 halamanRejilla Capitulo 8Wendy Paola Barragan JimenezBelum ada peringkat
- Replicación Del AdnDokumen28 halamanReplicación Del AdnisaacBelum ada peringkat
- Daño, Reparación y Recombinación Celular.Dokumen14 halamanDaño, Reparación y Recombinación Celular.yeson David urbanoBelum ada peringkat
- 3.trabajo ADN 4 MedioDokumen4 halaman3.trabajo ADN 4 Medioroxana_núñez_2Belum ada peringkat
- GENETICADokumen14 halamanGENETICAYANETHBelum ada peringkat
- 20 PolirribosomaDokumen1 halaman20 Polirribosomaapi-3700689Belum ada peringkat
- Sistema EndomembranosoDokumen121 halamanSistema EndomembranosoProfaRebecaManriqueBelum ada peringkat
- Herramientas de La Ingeniería GenéticaDokumen2 halamanHerramientas de La Ingeniería GenéticaMetzli RomanoBelum ada peringkat
- Unidad 1 y 2 - Mantenimiento y Variabilidad Del ADN NuclearDokumen7 halamanUnidad 1 y 2 - Mantenimiento y Variabilidad Del ADN NuclearEmma OchovaBelum ada peringkat
- Mecanismos de Reparación de Daños en La Molécula de Adn: Ángel Radamés Rábago MonzónDokumen11 halamanMecanismos de Reparación de Daños en La Molécula de Adn: Ángel Radamés Rábago MonzónRadamés RábagoBelum ada peringkat
- 11 FactTranscripEnfermedadesDokumen5 halaman11 FactTranscripEnfermedadesJenny TorresHBelum ada peringkat
- Ciencias ÓmicasDokumen3 halamanCiencias Ómicasandrea laraBelum ada peringkat
- 2 Microinyección de Adn en Pronúcleo o en Citoplasma de Cigoto (Óvulo Fecundado) y Microinyección Del Adn en Células EmbrionariasDokumen20 halaman2 Microinyección de Adn en Pronúcleo o en Citoplasma de Cigoto (Óvulo Fecundado) y Microinyección Del Adn en Células EmbrionariasMilenaBelum ada peringkat
- Ejercicio de ReplicaciónDokumen7 halamanEjercicio de ReplicaciónAngelesTorresBelum ada peringkat
- Kahoot - Flujo de InformacionDokumen12 halamanKahoot - Flujo de InformacioncrisBelum ada peringkat
- TALLER No 5 Replicacion ADNDokumen2 halamanTALLER No 5 Replicacion ADNKaterin GalvisBelum ada peringkat
- La Siguiente Es Una HEBRA DE ADN CODIFICANTE de ADNm BiologiaDokumen5 halamanLa Siguiente Es Una HEBRA DE ADN CODIFICANTE de ADNm BiologiaJhoan SerranoBelum ada peringkat
- Transmisión de La Información Genética PDFDokumen7 halamanTransmisión de La Información Genética PDFAndrez HernandezBelum ada peringkat
- Replicacion Alberts - En.esDokumen18 halamanReplicacion Alberts - En.esPaula NastarBelum ada peringkat
- Cuestionario Fredy Calizaya LlatasiDokumen3 halamanCuestionario Fredy Calizaya LlatasiFredy GrimaldoBelum ada peringkat
- CRISPR Cas9Dokumen8 halamanCRISPR Cas9Andrea OcampoBelum ada peringkat
- Los Marcadores Moleculares en El Mejoramiento Genético de La PDFDokumen13 halamanLos Marcadores Moleculares en El Mejoramiento Genético de La PDFivanBelum ada peringkat
- Regulación GénicaDokumen3 halamanRegulación GénicaFrancisco Javier F. LeónBelum ada peringkat
- Mock ExamDokumen7 halamanMock Examquique ddmBelum ada peringkat