Anda mungkin juga menyukai
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- All About MSDokumen18 halamanAll About MSMuhammad AkrimBelum ada peringkat
- Fever PDFDokumen2 halamanFever PDFMuhammad AkrimBelum ada peringkat
- Sympathy, Empathy, and Compassion: A Grounded Theory Study of Palliative Care Patients' Understandings, Experiences, and PreferencesDokumen11 halamanSympathy, Empathy, and Compassion: A Grounded Theory Study of Palliative Care Patients' Understandings, Experiences, and PreferencesMuhammad AkrimBelum ada peringkat
- 2.a Lesson in Empathy.16Dokumen2 halaman2.a Lesson in Empathy.16Muhammad AkrimBelum ada peringkat
- 3Dokumen5 halaman3Muhammad AkrimBelum ada peringkat
- Neonatal Care Pocket Guide For Hospital Physicians PDFDokumen319 halamanNeonatal Care Pocket Guide For Hospital Physicians PDFelmaadawy2002100% (7)
- Dapus Referat No.4Dokumen7 halamanDapus Referat No.4Muhammad AkrimBelum ada peringkat
- Dapus Referat No. 11Dokumen4 halamanDapus Referat No. 11Muhammad AkrimBelum ada peringkat
- Dapus Referat No.14Dokumen10 halamanDapus Referat No.14Muhammad AkrimBelum ada peringkat
- Dapus Referat No.12Dokumen8 halamanDapus Referat No.12Muhammad AkrimBelum ada peringkat
- Dapus Referat No.12Dokumen8 halamanDapus Referat No.12Muhammad AkrimBelum ada peringkat
- Nehig PDFDokumen10 halamanNehig PDFMuhammad AkrimBelum ada peringkat
- Predictive Validity of Four Nicotine Dependence Measures in A College SampleDokumen10 halamanPredictive Validity of Four Nicotine Dependence Measures in A College SampleMuhammad AkrimBelum ada peringkat
- Dapus Referat No.8Dokumen6 halamanDapus Referat No.8Muhammad AkrimBelum ada peringkat
- Alcohol Use DisordersDokumen11 halamanAlcohol Use DisordersMuhammad AkrimBelum ada peringkat
- Variants in Nicotinic Receptors and Risk For Nicotine DependenceDokumen9 halamanVariants in Nicotinic Receptors and Risk For Nicotine DependenceMuhammad AkrimBelum ada peringkat
- Dapus Referat No.17Dokumen9 halamanDapus Referat No.17Muhammad AkrimBelum ada peringkat
- September2014 - Vantilburg For WebsiteDokumen14 halamanSeptember2014 - Vantilburg For WebsiteMuhammad AkrimBelum ada peringkat
- Autobiographical Memory in Non-Amnesic Alcohol-Dependent PatientsDokumen9 halamanAutobiographical Memory in Non-Amnesic Alcohol-Dependent PatientsMuhammad AkrimBelum ada peringkat
- Dapus Referat No.9Dokumen13 halamanDapus Referat No.9Muhammad AkrimBelum ada peringkat
- Jurnal 9 PDFDokumen46 halamanJurnal 9 PDFMuhammad AkrimBelum ada peringkat
- FEAR OF HOLE Ade WibowoArif PDFDokumen19 halamanFEAR OF HOLE Ade WibowoArif PDFChrisBelum ada peringkat
- Jurnal 16Dokumen9 halamanJurnal 16Muhammad AkrimBelum ada peringkat
- Dapus Referat No.4Dokumen7 halamanDapus Referat No.4Muhammad AkrimBelum ada peringkat
- Validation PointsDokumen6 halamanValidation PointsMuhammad AkrimBelum ada peringkat
- 2.1 Chapter6Dokumen48 halaman2.1 Chapter6Muhammad AkrimBelum ada peringkat
- Rain Natomy: Adapted From Human Anatomy & Physiology by Marieb and Hoehn (9 Ed.)Dokumen16 halamanRain Natomy: Adapted From Human Anatomy & Physiology by Marieb and Hoehn (9 Ed.)vibs1994Belum ada peringkat
- YMEM5876 ProofDokumen11 halamanYMEM5876 ProofMuhammad AkrimBelum ada peringkat
- Perforated Appendicitis PDFDokumen8 halamanPerforated Appendicitis PDFMuhammad AkrimBelum ada peringkat
- Professional Med J Q 2014 21 1 27 32Dokumen6 halamanProfessional Med J Q 2014 21 1 27 32Pinandhito LatukolanBelum ada peringkat
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (120)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Cumulative NCLEX Test May 2023Dokumen29 halamanCumulative NCLEX Test May 2023Julie Ann100% (2)
- 22q11.2 GPC 2023Dokumen24 halaman22q11.2 GPC 2023Antoine NinoBelum ada peringkat
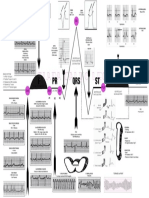
- Lesson Plan On EcgDokumen3 halamanLesson Plan On Ecganimesh panda100% (1)
- Diabetes and MassageDokumen23 halamanDiabetes and MassagemassagekevinBelum ada peringkat
- All Medical Syndromes 2022Dokumen141 halamanAll Medical Syndromes 2022Fercho MedBelum ada peringkat
- A Case Presentation of An Adult With Gestational Diabetes MellitusDokumen56 halamanA Case Presentation of An Adult With Gestational Diabetes MellitusAnn Mariz Dominguez100% (5)
- MedSurge HYPERCALCEMIADokumen9 halamanMedSurge HYPERCALCEMIAArfela Joy Dela CruzBelum ada peringkat
- Frog ALL SYSTEMS-1Dokumen85 halamanFrog ALL SYSTEMS-1Ranjeet PandeyBelum ada peringkat
- SBIL - Health QuestionnaireDokumen2 halamanSBIL - Health QuestionnaireKrishna GBBelum ada peringkat
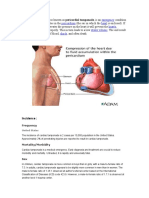
- Cardiac Tamponade, Also Known As Pericardial Tamponade, Is AnDokumen7 halamanCardiac Tamponade, Also Known As Pericardial Tamponade, Is AnSimran Josan100% (1)
- TMS ChecklistDokumen1 halamanTMS ChecklistMuhammad HussnainBelum ada peringkat
- Ventricular Isovolumetric Contractions: Tiara Calvo Leon, M.DDokumen1 halamanVentricular Isovolumetric Contractions: Tiara Calvo Leon, M.DSarah KhanBelum ada peringkat
- Ebook The Neuroicu Book PDF Full Chapter PDFDokumen67 halamanEbook The Neuroicu Book PDF Full Chapter PDFrobert.vann805100% (21)
- Krok 2 Medicine (EN) Attempt Review 1Dokumen52 halamanKrok 2 Medicine (EN) Attempt Review 1IS ZDBelum ada peringkat
- Citicoline Drug StudyDokumen2 halamanCiticoline Drug StudyJC LumayaBelum ada peringkat
- Biology: Pearson EdexcelDokumen32 halamanBiology: Pearson Edexcelbody fayez100% (1)
- Unknown 5Dokumen35 halamanUnknown 5Rayane NaegelsBelum ada peringkat
- Seminar On Fluid-and-Electrolyte-ImbalanceDokumen35 halamanSeminar On Fluid-and-Electrolyte-Imbalancemohamad dildarBelum ada peringkat
- JCM 12 01482 v2 PDFDokumen11 halamanJCM 12 01482 v2 PDFPutra FhonnaBelum ada peringkat
- Acute Respiratory Distress SyndromeDokumen3 halamanAcute Respiratory Distress Syndromekarenkaren09Belum ada peringkat
- 7 Urinary Disorders - 2012 - Small Animal Clinical Diagnosis by Laboratory Methods Fifth EditionDokumen30 halaman7 Urinary Disorders - 2012 - Small Animal Clinical Diagnosis by Laboratory Methods Fifth EditionNarvarte Hospital Veterinario de EspecialidadesBelum ada peringkat
- 3 MAJOR OPENINGS /right Atirum: Cardiovascular System Serious PericardiumDokumen3 halaman3 MAJOR OPENINGS /right Atirum: Cardiovascular System Serious PericardiumAebee Alcaraz100% (1)
- Inotropes and Vasoconstictor PackageDokumen25 halamanInotropes and Vasoconstictor PackageYoussef MokdadBelum ada peringkat
- ALS Manual 2010.version 1Dokumen84 halamanALS Manual 2010.version 1Elena How100% (1)
- AlgorithmACLS Tachycardia 200612Dokumen1 halamanAlgorithmACLS Tachycardia 200612YassarBelum ada peringkat
- Hyperthyroidism: - 60% Due To Grave's Disease - 40% Due To Miscellaneous CausesDokumen6 halamanHyperthyroidism: - 60% Due To Grave's Disease - 40% Due To Miscellaneous CausesLeitavia D.Belum ada peringkat
- Myocarditis & PericarditisDokumen28 halamanMyocarditis & PericarditisMahesh RathnayakeBelum ada peringkat
- NCP JenDokumen2 halamanNCP JenJenny Delos SantosBelum ada peringkat
- Hypoxic Ischemic Encephalopathy (Hie) : AlgorithmDokumen11 halamanHypoxic Ischemic Encephalopathy (Hie) : Algorithmsidharth sauravBelum ada peringkat
- Society For Vascular Surgery - What Is A Vascular Surgeon - 2020-06-25Dokumen6 halamanSociety For Vascular Surgery - What Is A Vascular Surgeon - 2020-06-25Talan DunnBelum ada peringkat