Anda mungkin juga menyukai
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (894)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (587)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (73)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Brain HerniationDokumen80 halamanBrain Herniationlamoleverde9297100% (1)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- Chest TubesDokumen3 halamanChest TubesKay BristolBelum ada peringkat
- Word Frequency - Based On 450 Million Word COCA CorpusDokumen89 halamanWord Frequency - Based On 450 Million Word COCA Corpusjoelson100% (2)
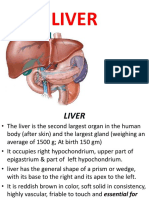
- LIVERDokumen18 halamanLIVERShivaniLeela100% (3)
- Mesenchymal CellsDokumen16 halamanMesenchymal Cellslamoleverde9297Belum ada peringkat
- Sex-Specific Patterns and Deregulation of Endocrine Pathways in The Gene Expression Profiles of Bangladeshi Adults Exposed To Arsenic Contaminated Drinking Water.Dokumen9 halamanSex-Specific Patterns and Deregulation of Endocrine Pathways in The Gene Expression Profiles of Bangladeshi Adults Exposed To Arsenic Contaminated Drinking Water.lamoleverde9297Belum ada peringkat
- Carcinogenesis TabacoDokumen17 halamanCarcinogenesis Tabacolamoleverde9297Belum ada peringkat
- Blank Disney ScheduleDokumen4 halamanBlank Disney Schedulelamoleverde9297Belum ada peringkat
- mrp1 ExpressionDokumen10 halamanmrp1 Expressionlamoleverde9297Belum ada peringkat
- Sex-Specific Patterns and Deregulation of Endocrine Pathways in The Gene Expression Profiles of Bangladeshi Adults Exposed To Arsenic Contaminated Drinking Water.Dokumen9 halamanSex-Specific Patterns and Deregulation of Endocrine Pathways in The Gene Expression Profiles of Bangladeshi Adults Exposed To Arsenic Contaminated Drinking Water.lamoleverde9297Belum ada peringkat
- GINA Pocket2013 May15Dokumen0 halamanGINA Pocket2013 May15David BarzalloBelum ada peringkat
- Bacteria PDFDokumen13 halamanBacteria PDFJohn Christopher LucesBelum ada peringkat
- AcromegaliaDokumen16 halamanAcromegalialamoleverde9297Belum ada peringkat
- Epidemiology of Lung Cancer ACCP Evidence-Based Clinical Practice Guidelines. 2007pdfDokumen27 halamanEpidemiology of Lung Cancer ACCP Evidence-Based Clinical Practice Guidelines. 2007pdflamoleverde9297Belum ada peringkat
- Arsenic and Cigarette Smoke Synergistically Increase DNA Oxidation in The Lung. - 2006 - Hays Et AlDokumen10 halamanArsenic and Cigarette Smoke Synergistically Increase DNA Oxidation in The Lung. - 2006 - Hays Et Allamoleverde9297Belum ada peringkat
- Organophosphate Poisoning GuidelineDokumen5 halamanOrganophosphate Poisoning Guidelinelamoleverde9297Belum ada peringkat
- TB 2Dokumen26 halamanTB 2lamoleverde9297Belum ada peringkat
- Surgery For Cerebral Contusions: Rationale and Practice: Review ArticleDokumen4 halamanSurgery For Cerebral Contusions: Rationale and Practice: Review ArticleIshan SharmaBelum ada peringkat
- The Homeopathic Treatment of LymeDokumen31 halamanThe Homeopathic Treatment of LymeMartoufi100% (2)
- Lavalvulatricuspide Esagrandesconocidajmrevuelta 111125055001 Phpalpp02Dokumen105 halamanLavalvulatricuspide Esagrandesconocidajmrevuelta 111125055001 Phpalpp02juan victor bejar sayaBelum ada peringkat
- Goat Farming 2004Dokumen154 halamanGoat Farming 2004Jahn Symon RiveraBelum ada peringkat
- Semen AnalysisDokumen3 halamanSemen Analysisameerabest80% (5)
- LAMPIRAN RET PELAYANAN KESEHATAN (Rev)Dokumen167 halamanLAMPIRAN RET PELAYANAN KESEHATAN (Rev)RafliBelum ada peringkat
- Gas Gangrene.Dokumen24 halamanGas Gangrene.Jared GreenBelum ada peringkat
- Diagnostic MethodsDokumen93 halamanDiagnostic MethodsSyukri La Ranti100% (3)
- ANTI OBESITY CAMPDokumen13 halamanANTI OBESITY CAMPgirishlambaBelum ada peringkat
- TABLE (Ascaris, Trichuris, and Enterobius)Dokumen3 halamanTABLE (Ascaris, Trichuris, and Enterobius)TRISHA MAE ORDONABelum ada peringkat
- Ambahan: Ambahans Inscribed On Plants in A Bamboo Grove (Source: Antoon Postma)Dokumen10 halamanAmbahan: Ambahans Inscribed On Plants in A Bamboo Grove (Source: Antoon Postma)Mernie Grace DionesioBelum ada peringkat
- Assisted Reproductive Technologies OverviewDokumen35 halamanAssisted Reproductive Technologies OverviewPraluki HerliawanBelum ada peringkat
- Mohamed Ali El Maroush PH.D of Prosthodontic & Maxillofacial (Ain Shams University)Dokumen38 halamanMohamed Ali El Maroush PH.D of Prosthodontic & Maxillofacial (Ain Shams University)Ňøoř ĄłhųdàBelum ada peringkat
- 2nd Periodical TestDokumen11 halaman2nd Periodical TestGiYaIbarbiaBelum ada peringkat
- Milk PresentationDokumen14 halamanMilk PresentationNicole_Packard_6907Belum ada peringkat
- Factors That Damage the Stomach LiningDokumen8 halamanFactors That Damage the Stomach LiningTania Louise Pioquinto AbuanBelum ada peringkat
- Principles of Spine Trauma and Spinal Deformities PDFDokumen34 halamanPrinciples of Spine Trauma and Spinal Deformities PDFVirlan Vasile CatalinBelum ada peringkat
- Annotated Reading Dengue FeverDokumen1 halamanAnnotated Reading Dengue FeverGoRbz SarmientoBelum ada peringkat
- Bone TissueDokumen66 halamanBone TissueFunikawati Baldiyah100% (1)
- Care Sheet - Axolotl (Ambystoma Mexicanum)Dokumen3 halamanCare Sheet - Axolotl (Ambystoma Mexicanum)John GamesbyBelum ada peringkat
- Alveolar Osteitis A Comprehensive Review ofDokumen10 halamanAlveolar Osteitis A Comprehensive Review ofFakhrul ImamBelum ada peringkat
- Staying Healthy as a StudentDokumen19 halamanStaying Healthy as a StudentRachma Susteriana PutriBelum ada peringkat
- Cellulitis: Clinical Review: Be The First To CommentDokumen9 halamanCellulitis: Clinical Review: Be The First To CommentAnonymous 1nMTZWmzBelum ada peringkat
- CRI Annual Report 2006Dokumen10 halamanCRI Annual Report 2006aptureincBelum ada peringkat
- Capybara SchoolprojectDokumen3 halamanCapybara SchoolprojectTom HoldaBelum ada peringkat
- Viral Diseases and the Immune SystemDokumen27 halamanViral Diseases and the Immune SystemHendika Ariyo SBelum ada peringkat
- CEVA ISAP2008 Sassi Et AlDokumen48 halamanCEVA ISAP2008 Sassi Et AlJef SassiBelum ada peringkat