Anda mungkin juga menyukai
- Use of Chitosan-Modified Bentonite For Removal of Cu2+, CL and 2,4-Dichlorophenoxyacetic Acid (2,4-D) From Aqueous SolutionDokumen6 halamanUse of Chitosan-Modified Bentonite For Removal of Cu2+, CL and 2,4-Dichlorophenoxyacetic Acid (2,4-D) From Aqueous Solutionmisterno2Belum ada peringkat
- The Role of Renewable Energy Sources On Electricity Prices in Spain. A Maximum Entropy Econometric ModelDokumen11 halamanThe Role of Renewable Energy Sources On Electricity Prices in Spain. A Maximum Entropy Econometric Modelmisterno2Belum ada peringkat
- Phase Equilibria of Alkali Nitrates (NaNO3, KNO3, RbNO3, CsNO3) in Glycerol + H2O System at Various TemperaturesDokumen8 halamanPhase Equilibria of Alkali Nitrates (NaNO3, KNO3, RbNO3, CsNO3) in Glycerol + H2O System at Various Temperaturesmisterno2Belum ada peringkat
- Developing A Steady-State Kinetic Model For Industrial Scale Semi-Regenerative Catalytic Naphtha Reforming ProcessDokumen6 halamanDeveloping A Steady-State Kinetic Model For Industrial Scale Semi-Regenerative Catalytic Naphtha Reforming Processmisterno2Belum ada peringkat
- Integrated Planning of Electricity, Gas and Heat Supply To MunicipalityDokumen18 halamanIntegrated Planning of Electricity, Gas and Heat Supply To Municipalitymisterno2Belum ada peringkat
- Corrosion Education As A ToolDokumen14 halamanCorrosion Education As A Toolmisterno2Belum ada peringkat
- Effects of Nano-Sized Titanium Powder On The Anti-Corrosion Property of Epoxy Coatings On SteelDokumen6 halamanEffects of Nano-Sized Titanium Powder On The Anti-Corrosion Property of Epoxy Coatings On Steelmisterno2Belum ada peringkat
- Performance Based Evaluation of Industrial Grade Resins Duolite ARA-9366 and Duolite A-368Dokumen8 halamanPerformance Based Evaluation of Industrial Grade Resins Duolite ARA-9366 and Duolite A-368misterno2Belum ada peringkat
- Phase Equilibrium of NaNO3/KNO3 in Glycerol + H2O Mixed Solvent System at 288.15 K and 298.15 KDokumen10 halamanPhase Equilibrium of NaNO3/KNO3 in Glycerol + H2O Mixed Solvent System at 288.15 K and 298.15 Kmisterno2Belum ada peringkat
- Velocity Profile Simulation For Natural Gasy EngDokumen5 halamanVelocity Profile Simulation For Natural Gasy Engmisterno2Belum ada peringkat
- Crystallization Kinetics of Precipitating Calcium From Yellow Phosphorus Slag LixiviumDokumen5 halamanCrystallization Kinetics of Precipitating Calcium From Yellow Phosphorus Slag Lixiviummisterno2Belum ada peringkat
- Microstructure and Physical-Chemical Properties of Petroleum Coke As CarburizerDokumen4 halamanMicrostructure and Physical-Chemical Properties of Petroleum Coke As Carburizermisterno2Belum ada peringkat
- Corrosion Mechanism of Cemented Soil in MgCl2 SolutionDokumen6 halamanCorrosion Mechanism of Cemented Soil in MgCl2 Solutionmisterno2Belum ada peringkat
- Optimization of Total Flavonoids Extraction From Coreopsis Tinctoria Nutt.Dokumen6 halamanOptimization of Total Flavonoids Extraction From Coreopsis Tinctoria Nutt.misterno2Belum ada peringkat
- Malvic EngDokumen7 halamanMalvic Engmisterno2Belum ada peringkat
- Croatia's Energy Sector Development by 2050 and CO2 Emission ReductionsDokumen7 halamanCroatia's Energy Sector Development by 2050 and CO2 Emission Reductionsmisterno2Belum ada peringkat
- Corrosion Inhibition of Oil-Well Steel (N-80) in Simulated Hydrogen Sulphide Environment by Ferrous Gluconate and Synthetic MagnetiteDokumen8 halamanCorrosion Inhibition of Oil-Well Steel (N-80) in Simulated Hydrogen Sulphide Environment by Ferrous Gluconate and Synthetic Magnetitemisterno2Belum ada peringkat
- Health, Safety and Environment Policy of Crosco, Integrated Drilling & Well Services Co., Ltd. - Application of HSE Practice in Onshore DrillingDokumen4 halamanHealth, Safety and Environment Policy of Crosco, Integrated Drilling & Well Services Co., Ltd. - Application of HSE Practice in Onshore Drillingmisterno2Belum ada peringkat
- Effect of Ph-Values On The Contact Angle and Interfacial TensionDokumen6 halamanEffect of Ph-Values On The Contact Angle and Interfacial Tensionmisterno2Belum ada peringkat
- Comparative Analysis of The Efficacies of Ferrous Gluconate and Synthetic Magnetite As Sulphide Scavengers in Oil and Gas Drilling OperationsDokumen6 halamanComparative Analysis of The Efficacies of Ferrous Gluconate and Synthetic Magnetite As Sulphide Scavengers in Oil and Gas Drilling Operationsmisterno2Belum ada peringkat
- Bioethanol Production and Power Generation For Co-ProductionDokumen7 halamanBioethanol Production and Power Generation For Co-Productionmisterno2Belum ada peringkat
- Analysis of Geothermal Energy Exploitation On Karlovac Geothermal FieldDokumen7 halamanAnalysis of Geothermal Energy Exploitation On Karlovac Geothermal Fieldmisterno2Belum ada peringkat
- Mapping of The Late Miocene Sandstone Facies Using Indicator KrigingDokumen9 halamanMapping of The Late Miocene Sandstone Facies Using Indicator Krigingmisterno2Belum ada peringkat
- Neural Network Prediction of Porosity and Permeability of Heterogeneous Gas Sand Reservoirs Using NMR and Conventional LogsDokumen10 halamanNeural Network Prediction of Porosity and Permeability of Heterogeneous Gas Sand Reservoirs Using NMR and Conventional Logsmisterno2Belum ada peringkat
- How To Address The Challanges of The Climate PreservationDokumen8 halamanHow To Address The Challanges of The Climate Preservationmisterno2Belum ada peringkat
- Corrosion Education As A ToolDokumen14 halamanCorrosion Education As A Toolmisterno2Belum ada peringkat
- Confusion of Forecasts (And) Forecasts of Confusion in Energy SectorDokumen6 halamanConfusion of Forecasts (And) Forecasts of Confusion in Energy Sectormisterno2Belum ada peringkat
- Zamanian Eng PDFDokumen8 halamanZamanian Eng PDFmisterno2Belum ada peringkat
- Development of An Artificial Neural Network Model To Predict CO2 Minimum Miscibility PressureDokumen4 halamanDevelopment of An Artificial Neural Network Model To Predict CO2 Minimum Miscibility Pressuremisterno2Belum ada peringkat
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (587)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (73)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (265)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (119)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Biobutanol - Advantages and Disavantages. Comparision To Ethanol.Dokumen9 halamanBiobutanol - Advantages and Disavantages. Comparision To Ethanol.Phi TiêuBelum ada peringkat
- Indonesia Gearing Towards Bio-Ethanol - Nov08Dokumen29 halamanIndonesia Gearing Towards Bio-Ethanol - Nov08Steve Lee100% (1)
- Exam 32007 Form 2Dokumen5 halamanExam 32007 Form 2Wiam BaallaBelum ada peringkat
- Ethanol From Banana Peels Investigatory ProjectDokumen5 halamanEthanol From Banana Peels Investigatory Projectsapphiredeguzman76% (17)
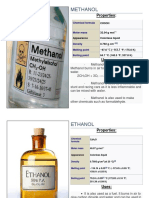
- Methanol: PropertiesDokumen5 halamanMethanol: PropertiesAlaidja BayonoBelum ada peringkat
- Question: Consider A Closed Vessel Initially Containing 1 Mol of Pure EthylDokumen2 halamanQuestion: Consider A Closed Vessel Initially Containing 1 Mol of Pure EthylJhowelaine AbrilBelum ada peringkat
- APES Types of Energy TableDokumen5 halamanAPES Types of Energy TableirregularflowersBelum ada peringkat
- Plant Design ProjectDokumen8 halamanPlant Design ProjectAbhishek SagarBelum ada peringkat
- Previous HSE Questions From The Chapter "Alcohols, Phenols and Ethers"Dokumen3 halamanPrevious HSE Questions From The Chapter "Alcohols, Phenols and Ethers"Nikhil MathewBelum ada peringkat
- Microbes in Human WelfareDokumen12 halamanMicrobes in Human WelfareDr.kaleem AhmedBelum ada peringkat
- Silica Sulfuric Acid An Efficient and Reusable Catalyst For The One-Pot Synthesis of 3,4-Dihydropyrimidin-2 (1H) - OnesDokumen3 halamanSilica Sulfuric Acid An Efficient and Reusable Catalyst For The One-Pot Synthesis of 3,4-Dihydropyrimidin-2 (1H) - OnesDuy Phuc LeBelum ada peringkat
- United States Patent (19) : Assignee: National Distillers and ChemicalDokumen10 halamanUnited States Patent (19) : Assignee: National Distillers and ChemicalantavenaBelum ada peringkat
- Determination of Alcohol Content in Alcoholic BeveragesDokumen7 halamanDetermination of Alcohol Content in Alcoholic BeveragesKaye Danielle HilomenBelum ada peringkat
- Carotenuto Giuseppina PDFDokumen459 halamanCarotenuto Giuseppina PDFGonca TuncayBelum ada peringkat
- Optimization of An Industrial Medium From Molasses For Bioethanol Production Using The Taguchi Statistical Experimental-Design MethodDokumen9 halamanOptimization of An Industrial Medium From Molasses For Bioethanol Production Using The Taguchi Statistical Experimental-Design MethodSagar ChaudharyBelum ada peringkat
- 1,2-Propylene glycol technical leafletDokumen8 halaman1,2-Propylene glycol technical leafletMurat ÇiniBelum ada peringkat
- Chapter 3 and 4 (TBD)Dokumen6 halamanChapter 3 and 4 (TBD)tekhiesBelum ada peringkat
- Miper Kurnool Organic ChemistryDokumen83 halamanMiper Kurnool Organic ChemistryNaeem RehmanBelum ada peringkat
- Formula Mass and Mole ConceptDokumen6 halamanFormula Mass and Mole ConceptGottfried Wilhelm LeibnizBelum ada peringkat
- State Excise ManualDokumen106 halamanState Excise Manualajeez86100% (1)
- Fermentation of Glucose Using YeastDokumen3 halamanFermentation of Glucose Using YeastBoomdayBelum ada peringkat
- Alcohols and PhenolsDokumen11 halamanAlcohols and PhenolsRovic MelladoBelum ada peringkat
- Experiment 3 - Carboxylic Acid and DerivativesDokumen3 halamanExperiment 3 - Carboxylic Acid and DerivativesFaris SyahmiBelum ada peringkat
- Prosses Validation Protocol For EnrofolxacineDokumen22 halamanProsses Validation Protocol For Enrofolxacineمحمد عطاBelum ada peringkat
- MLS121L02 CompoundidphyspropDokumen1 halamanMLS121L02 CompoundidphyspropTrisha CabusaoBelum ada peringkat
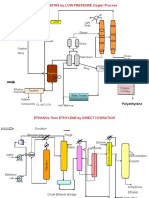
- POLY-OLEFINS by LOW PRESSURE Ziegler Process: Aluminum Alkyl Drier Light Ends DiluentDokumen3 halamanPOLY-OLEFINS by LOW PRESSURE Ziegler Process: Aluminum Alkyl Drier Light Ends Diluentessakkiraj.mBelum ada peringkat
- A Practical Handbook On TheDokumen145 halamanA Practical Handbook On TheGregoire De IzarraBelum ada peringkat
- Tecumseh OV195EA - OV195XA Engine ManualDokumen32 halamanTecumseh OV195EA - OV195XA Engine Manual"Rufus"0% (1)
- Production of Ethanol From MolassesDokumen13 halamanProduction of Ethanol From Molassesvivek gangwar80% (15)
- 7404 2 QP Chemistry AS 23may23 AMDokumen32 halaman7404 2 QP Chemistry AS 23may23 AMsai vanamBelum ada peringkat