Anda mungkin juga menyukai
- Diapositivas de EstafilococosDokumen33 halamanDiapositivas de EstafilococosMilagrOs Benites Reyes100% (1)
- Sistema CardiovascularDokumen98 halamanSistema CardiovascularIncy LopezBelum ada peringkat
- 13 Hemostasia-1Dokumen55 halaman13 Hemostasia-1Jessica ARBelum ada peringkat
- Banco de Preguntas Segundo Parcial LourdesDokumen7 halamanBanco de Preguntas Segundo Parcial LourdesMartínez Katerin BrigithBelum ada peringkat
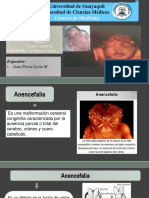
- Casoclinico AnencefaliaDokumen9 halamanCasoclinico AnencefaliaJesus Smaikel Teran MuenalaBelum ada peringkat
- Practica .LipoproteinasDokumen10 halamanPractica .LipoproteinasKarla Liszet Gonzalez HernandezBelum ada peringkat
- HemotoraxDokumen4 halamanHemotoraxVicente MartínezBelum ada peringkat
- Citotoxicidad celular: CTL, células NK y sus receptoresDokumen13 halamanCitotoxicidad celular: CTL, células NK y sus receptoresRaquel GuevaraBelum ada peringkat
- Metabolismo Del ARNDokumen53 halamanMetabolismo Del ARNJuly PachecoBelum ada peringkat
- Final SQX 2021Dokumen14 halamanFinal SQX 2021Anny JiménezBelum ada peringkat
- Práctica 1 Química SanguíneaDokumen14 halamanPráctica 1 Química SanguíneaSandy MoralesBelum ada peringkat
- Generalidades Del NúcleoDokumen5 halamanGeneralidades Del NúcleoCarlo CepulvedaBelum ada peringkat
- Patologia de La Cirrosis HepaticaDokumen13 halamanPatologia de La Cirrosis HepaticaLiliana Portal VillegasBelum ada peringkat
- CCL AlitiasicaDokumen6 halamanCCL AlitiasicaAlexis CarvajalBelum ada peringkat
- 3 Fisiologia UrologicaDokumen62 halaman3 Fisiologia UrologicaKarina Alexandra0% (1)
- Analisis Clinicos Por TemasDokumen414 halamanAnalisis Clinicos Por TemasEduardo Romero100% (1)
- Trauma de ToraxDokumen36 halamanTrauma de ToraxTennessee Ayala MendozaBelum ada peringkat
- Anatomia Del Sistema CardiacoDokumen8 halamanAnatomia Del Sistema CardiacoLoren RamirezBelum ada peringkat
- Hemostasia, Formación de Trombo y Proceso de CoagulaciónDokumen3 halamanHemostasia, Formación de Trombo y Proceso de CoagulaciónPaulyMartinezBelum ada peringkat
- Generalidades y Bases de OncologíaDokumen5 halamanGeneralidades y Bases de Oncologíaisidora canalesBelum ada peringkat
- Hemoglobina FetalDokumen3 halamanHemoglobina FetalPayola TitoBelum ada peringkat
- OntogeniaDokumen57 halamanOntogeniaEmilio Davila Alvarez100% (1)
- Enfermedades Linfoproliferativas PDFDokumen43 halamanEnfermedades Linfoproliferativas PDFGraham Paredes100% (1)
- Catedra de OncologiaDokumen49 halamanCatedra de OncologiaUSB_90Belum ada peringkat
- Patologias de LaringeDokumen14 halamanPatologias de LaringeOrlando SepulvedaBelum ada peringkat
- Cavidad Abdominal y AbDokumen31 halamanCavidad Abdominal y Abchampfiguereo2753Belum ada peringkat
- Tarea Final Hematologia (Autoguardado)Dokumen23 halamanTarea Final Hematologia (Autoguardado)Cindy MassielBelum ada peringkat
- ProlactinemiaDokumen15 halamanProlactinemiaMyriam MendozaBelum ada peringkat
- Protocolo Post-Operatoria #5Dokumen2 halamanProtocolo Post-Operatoria #5Joseph Antonio A. GomezBelum ada peringkat
- Control Del Ciclo Celular y Reparación Del ADNDokumen24 halamanControl Del Ciclo Celular y Reparación Del ADNRafa GomezBelum ada peringkat
- INJURIA CELULAR Dra JaraDokumen57 halamanINJURIA CELULAR Dra JaraEdward ChacmaBelum ada peringkat
- Sindrome de Ehlers-DanlosDokumen29 halamanSindrome de Ehlers-DanlosJefMorenoBelum ada peringkat
- Tipos de Células ProstáticasDokumen7 halamanTipos de Células ProstáticasKevin Urbina DiazBelum ada peringkat
- Enfermedad Pulmonar Intersticial DisfusaDokumen10 halamanEnfermedad Pulmonar Intersticial DisfusadrheayBelum ada peringkat
- Cardiopatías congénitas no cianóticasDokumen46 halamanCardiopatías congénitas no cianóticasEidibet Navarro100% (1)
- Radiografia de AbdomenDokumen69 halamanRadiografia de Abdomencheskubee1100% (2)
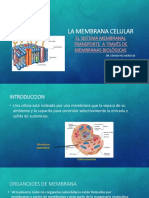
- Membrana CelularDokumen22 halamanMembrana CelularManuel Cespedes100% (1)
- Anatomia y Semiologia Del CuelloDokumen26 halamanAnatomia y Semiologia Del CuellokatyyokisBelum ada peringkat
- Tumores cardiacos más frecuentesDokumen69 halamanTumores cardiacos más frecuentesPau HcBelum ada peringkat
- Membrana Celular Presentacion PDFDokumen73 halamanMembrana Celular Presentacion PDFSergio Paz MeruviaBelum ada peringkat
- Cuestionario Fisiopatología - Diabetes MellitusDokumen7 halamanCuestionario Fisiopatología - Diabetes MellitusCarla Marliz Ore Jaime100% (1)
- Pericarditis AgudaDokumen27 halamanPericarditis AgudaCesar Giovanni Cordova100% (1)
- Metabolismo Del HierroDokumen17 halamanMetabolismo Del HierroLindsay Cuadros Leon0% (1)
- Sangrado Del Primer TrimestreDokumen42 halamanSangrado Del Primer TrimestreveguetaBelum ada peringkat
- Universidad Nacional José Faustino Sánchez CarriónDokumen31 halamanUniversidad Nacional José Faustino Sánchez CarriónIris Rộxậnậ Rậmộs CHậhuậ100% (1)
- Neutrófilo GingivalDokumen14 halamanNeutrófilo GingivalMelissa Mariví CastellanosBelum ada peringkat
- EpistaxisDokumen9 halamanEpistaxisMartín BonillaBelum ada peringkat
- Tumores PulmonaresDokumen21 halamanTumores PulmonaresAlejandra Sime CoronelBelum ada peringkat
- Función EndotelialDokumen35 halamanFunción EndotelialAngel Mauricio Cardenas Pineda100% (1)
- VPHDokumen10 halamanVPHmonyckgcBelum ada peringkat
- Temario Mundo Enarm 2019Dokumen15 halamanTemario Mundo Enarm 2019Alan LariosBelum ada peringkat
- ENZIMASDokumen51 halamanENZIMASDaniela Zuleta OvalleBelum ada peringkat
- NeutrofilosDokumen25 halamanNeutrofilosLuisa BernalBelum ada peringkat
- DAP y CIA en recién nacidosDokumen20 halamanDAP y CIA en recién nacidosMaria Yamilet Gonzalez LopezBelum ada peringkat
- Resumen VenodiseccionDokumen2 halamanResumen VenodiseccionAna KarenBelum ada peringkat
- Cáncer Pancreático ExocrinoDokumen32 halamanCáncer Pancreático ExocrinoEstefany MejiaBelum ada peringkat
- Mutaciones MurrayDokumen28 halamanMutaciones MurrayAdolfo Antonio Ríos AlcortaBelum ada peringkat
- Malformaciones Congénitas Del Cuello.Dokumen6 halamanMalformaciones Congénitas Del Cuello.Sebastian Gutierrez CastilloBelum ada peringkat
- Dimeros D Analisis de Sangre 2686 Mvi540Dokumen2 halamanDimeros D Analisis de Sangre 2686 Mvi540thorin8100% (1)
- Fiebre AmarillaDokumen13 halamanFiebre AmarillaArturo RecillasBelum ada peringkat
- AMEBIASISDokumen4 halamanAMEBIASISArturo RecillasBelum ada peringkat
- Características, Componentes y Anormalidades de La OrinaDokumen9 halamanCaracterísticas, Componentes y Anormalidades de La OrinaArturo Recillas100% (3)
- Glucólisis y Ciclo Del Acido CitricoDokumen3 halamanGlucólisis y Ciclo Del Acido CitricoArturo RecillasBelum ada peringkat
- COULOMBIMETRIADokumen4 halamanCOULOMBIMETRIAArturo RecillasBelum ada peringkat
- Evaluación de La Contaminación de Suelos Por Metales Pesados en Zonas Urbanas de Monterrey, Nuevo LeónDokumen7 halamanEvaluación de La Contaminación de Suelos Por Metales Pesados en Zonas Urbanas de Monterrey, Nuevo LeónArturo RecillasBelum ada peringkat
- Fenomeno SwarmingDokumen8 halamanFenomeno SwarmingArturo RecillasBelum ada peringkat
- Taller Final MecanicaDokumen3 halamanTaller Final MecanicaMARIA ALEJANDRA GONZALEZ BONILLABelum ada peringkat
- Tema 7 InteractomicaDokumen184 halamanTema 7 InteractomicaSoniaGarrichuBelum ada peringkat
- Libro Cap 1 y 2Dokumen19 halamanLibro Cap 1 y 2Jon SalazarBelum ada peringkat
- TEMA 2aDokumen39 halamanTEMA 2aJaime SnchzBelum ada peringkat
- Guía 1 (Bases de Datos Biológicas - Completo) 2023Dokumen22 halamanGuía 1 (Bases de Datos Biológicas - Completo) 2023MARIA BELEN BALTA BLANCOBelum ada peringkat
- Taller Tercer Corte BioquimicaDokumen3 halamanTaller Tercer Corte BioquimicaMARIA ALEJANDRA GONZALEZ BONILLABelum ada peringkat
- La Función de Los GPCR en La Percepción SensorialDokumen30 halamanLa Función de Los GPCR en La Percepción SensorialArturo Recillas100% (1)
- Dokument - 34 (1) .TRADUCIDODokumen132 halamanDokument - 34 (1) .TRADUCIDOJosé Ernesto Hugo QuispeBelum ada peringkat
- Análisis de la calidad de un bombón de chocolateDokumen13 halamanAnálisis de la calidad de un bombón de chocolateeddyguilty88100% (1)
- Inmunohistoquímica: Bloqueos y Artefactos en IHQDokumen12 halamanInmunohistoquímica: Bloqueos y Artefactos en IHQFranciscaBelum ada peringkat
- Métodos de Detección FluorométricaDokumen9 halamanMétodos de Detección FluorométricaMario RodríguezBelum ada peringkat
- Análisis ProteómicosDokumen39 halamanAnálisis ProteómicosArlette OsorioBelum ada peringkat