Anda mungkin juga menyukai
- A 05 - MP 6â°aula - ReflorestamentoDokumen55 halamanA 05 - MP 6â°aula - ReflorestamentoBianca ProençaBelum ada peringkat
- Pinhão-Roxo - Jatropha Gossypiifolia L. - Ervas Medicinais - Ficha Completa IlustradaDokumen9 halamanPinhão-Roxo - Jatropha Gossypiifolia L. - Ervas Medicinais - Ficha Completa IlustradaViam prudentiaeBelum ada peringkat
- 1 Lista de Exercícios (Estudo Dirigido 1 Avaliação)Dokumen4 halaman1 Lista de Exercícios (Estudo Dirigido 1 Avaliação)Renata CamposBelum ada peringkat
- CNatureza PlantasDokumen4 halamanCNatureza Plantasprof.vbatista100% (12)
- Morfologia de Folhas IFMADokumen12 halamanMorfologia de Folhas IFMAHicoxxBelum ada peringkat
- Ceiba SpeciosaDokumen5 halamanCeiba SpeciosaHugo HenriqueBelum ada peringkat
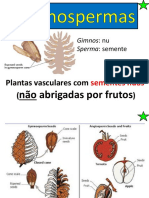
- Gimnospermas: principais grupos e reproduçãoDokumen24 halamanGimnospermas: principais grupos e reproduçãoWilliam SchultzBelum ada peringkat
- Aula 11. Grupos de Doenas de Plantas - Parte Inicial - Grupos I e II - 21-01-20Dokumen53 halamanAula 11. Grupos de Doenas de Plantas - Parte Inicial - Grupos I e II - 21-01-20Marcella Gomes EstevesBelum ada peringkat
- XilemaDokumen1 halamanXilemavhuppes92Belum ada peringkat
- Angiospermas: Plantas com Flores e FrutosDokumen4 halamanAngiospermas: Plantas com Flores e FrutosSaraiva JuniorBelum ada peringkat
- Reprodução e multiplicação de plantasDokumen57 halamanReprodução e multiplicação de plantasMeghan WilsonBelum ada peringkat
- Ecossistema natureza dicionárioDokumen79 halamanEcossistema natureza dicionárioJaquelineMouraBelum ada peringkat
- Cultivo do Feijão MacassarDokumen76 halamanCultivo do Feijão MacassarAnnykellyvBelum ada peringkat
- Manchas Foliares, Míldios, Oídios, FerrugemDokumen40 halamanManchas Foliares, Míldios, Oídios, Ferrugemjrturra100% (1)
- ATIVDokumen3 halamanATIVRACHEL PEREIRABelum ada peringkat
- Biologia - Morfologia das angiospermasDokumen2 halamanBiologia - Morfologia das angiospermasbobgilpordosol2857Belum ada peringkat
- Farmacobotânica UFSJDokumen4 halamanFarmacobotânica UFSJHermínio BenítezBelum ada peringkat
- Cultura Da CebolaDokumen42 halamanCultura Da CebolaDANILO MESSIAS DE OLIVEIRABelum ada peringkat
- Guia de Cultivo de Rosas Do DesertoDokumen3 halamanGuia de Cultivo de Rosas Do DesertoMarcelo DavidBelum ada peringkat
- RoteiroDokumen19 halamanRoteirooṣó igboBelum ada peringkat
- Atividades Reino PlantaeDokumen1 halamanAtividades Reino PlantaeDaucileliBelum ada peringkat
- Aula 1 Hort 2018Dokumen122 halamanAula 1 Hort 2018Ramon Rêgo100% (1)
- Estudo Taxonômico Do Gênero Cuphea P.Browne (Lythraceae) No Rio Grande Do Sul, BrasilDokumen163 halamanEstudo Taxonômico Do Gênero Cuphea P.Browne (Lythraceae) No Rio Grande Do Sul, BrasilRodrigo SouzaBelum ada peringkat
- Distribuicao e Abundancia de Especies Arboreas Do Cerrado Sensu Stricto No Pantanal, Mato Grosso, Brasil PDFDokumen8 halamanDistribuicao e Abundancia de Especies Arboreas Do Cerrado Sensu Stricto No Pantanal, Mato Grosso, Brasil PDFTravis VasquezBelum ada peringkat
- Exercicios Raiz Caule FolhasDokumen6 halamanExercicios Raiz Caule FolhasLarissa AndradeBelum ada peringkat
- Reino PlantaeDokumen21 halamanReino PlantaeBruno FelipeBelum ada peringkat
- Vitória RégiaDokumen4 halamanVitória RégiaJulio LealBelum ada peringkat
- Aula - Espécies Arboreas PDFDokumen44 halamanAula - Espécies Arboreas PDFNatu RebaBelum ada peringkat
- Fotomorfogênese em plantas: a ação da luz como sinal ambientalDokumen41 halamanFotomorfogênese em plantas: a ação da luz como sinal ambientalAugusto SimãoBelum ada peringkat
- Especialidade de CactosDokumen8 halamanEspecialidade de CactosCrycielle OlimpioBelum ada peringkat