Anda mungkin juga menyukai
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- T: G A P: Ables Roups of Cupuncture OintsDokumen6 halamanT: G A P: Ables Roups of Cupuncture OintsrikymediaBelum ada peringkat
- DedeDokumen8 halamanDedeHannah V. LacambraBelum ada peringkat
- Detailed Lesson PlanDokumen4 halamanDetailed Lesson PlanJudith CabisoBelum ada peringkat
- Dog GoldendoodleDokumen8 halamanDog GoldendoodlejustasBelum ada peringkat
- Spermatogenesis and FertilizationDokumen21 halamanSpermatogenesis and FertilizationAyen FornollesBelum ada peringkat
- Saber Toothed CatDokumen4 halamanSaber Toothed CatMarie WilkersonBelum ada peringkat
- PM Artifacts WEBDokumen20 halamanPM Artifacts WEBSalman MemonBelum ada peringkat
- Red Dog by Louis de Bernières Sample ChapterDokumen12 halamanRed Dog by Louis de Bernières Sample ChapterRandomHouseAU83% (6)
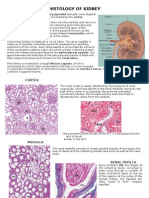
- Histology of KidneyDokumen4 halamanHistology of KidneyIzzi FekratBelum ada peringkat
- Robotics in Meat IndustryDokumen40 halamanRobotics in Meat IndustryDr Sarvadnya GhongdeBelum ada peringkat
- 7 Reading Task 1. For Questions 1-6, Read The Text Below and Decide Which Answer (A, B, C or D) Best Fits Each Gap. Fill in The Table Below The TextDokumen2 halaman7 Reading Task 1. For Questions 1-6, Read The Text Below and Decide Which Answer (A, B, C or D) Best Fits Each Gap. Fill in The Table Below The TextСофия ТимофееваBelum ada peringkat
- AQA Biology Seperate Science (Unit3) Summary NotesDokumen28 halamanAQA Biology Seperate Science (Unit3) Summary NotesKeval VaghjianiBelum ada peringkat
- Natural - HMS1 Ebook PDFDokumen120 halamanNatural - HMS1 Ebook PDFColoredCoded100% (2)
- PDFDokumen3 halamanPDFAnonymous tZpZgQBelum ada peringkat
- Descartes Pineal GlandDokumen20 halamanDescartes Pineal GlandKhaliq Dawood BuxBelum ada peringkat
- Lecture Outline: See Separate Powerpoint Slides For All Figures and Tables Pre-Inserted Into Powerpoint Without NotesDokumen61 halamanLecture Outline: See Separate Powerpoint Slides For All Figures and Tables Pre-Inserted Into Powerpoint Without NotesJharaBelum ada peringkat
- Pengaruh Umur Induk Dan Lama Penyimpanan Telur Terhadap Bobot Telur, Daya Tetas Dan Bobot Tetas Ayam Kedu Jengger MerahDokumen4 halamanPengaruh Umur Induk Dan Lama Penyimpanan Telur Terhadap Bobot Telur, Daya Tetas Dan Bobot Tetas Ayam Kedu Jengger MerahKKN-T Gorontalo 2020Belum ada peringkat
- Hotel Taj Palace PresentationDokumen33 halamanHotel Taj Palace PresentationBhupesh KumarBelum ada peringkat
- Dimensions of The DentureDokumen8 halamanDimensions of The DentureetequetaquiBelum ada peringkat
- Acute Flaccid Paralysis Surveillance PDFDokumen1 halamanAcute Flaccid Paralysis Surveillance PDFMutiara DewiBelum ada peringkat
- Whats The SettingDokumen2 halamanWhats The Settingapi-2428046300% (2)
- Personal IdentificationDokumen122 halamanPersonal Identificationkarl luisBelum ada peringkat
- 2013 ENDOCRINE PHARMACOLOGY Word Notes PDFDokumen52 halaman2013 ENDOCRINE PHARMACOLOGY Word Notes PDFNicole Opao100% (4)
- Worm Farming With MealwormsDokumen1 halamanWorm Farming With MealwormsVenketeshwar SwamiBelum ada peringkat
- Contractor Injury Report FormDokumen2 halamanContractor Injury Report Formmohamed11111981100% (1)
- Dual CementDokumen33 halamanDual CementStephani CastilloBelum ada peringkat
- Posterior Abdominal WallDokumen125 halamanPosterior Abdominal Wallarjun.k5796Belum ada peringkat
- Regulation of Cardiac Output and Venous Return by Dr. RoomiDokumen44 halamanRegulation of Cardiac Output and Venous Return by Dr. RoomiMudassar RoomiBelum ada peringkat
- STD 12 English Board Question Paper Maharashtra BoardDokumen22 halamanSTD 12 English Board Question Paper Maharashtra BoardTashvi KulkarniBelum ada peringkat
- Project Report On Indian BirdsDokumen9 halamanProject Report On Indian BirdsMridul kejriwal50% (2)