Anda mungkin juga menyukai
- Dissection of The Genetic Basis of Heterosis in An Elite Maize Hybrid by QTL Mapping in An Immortalized F2 PopulationDokumen8 halamanDissection of The Genetic Basis of Heterosis in An Elite Maize Hybrid by QTL Mapping in An Immortalized F2 PopulationvahidvasharBelum ada peringkat
- The Promising Corn-2009Dokumen1 halamanThe Promising Corn-2009vahidvasharBelum ada peringkat
- Positional Cloning of The Wheat VernalizationDokumen6 halamanPositional Cloning of The Wheat VernalizationvahidvasharBelum ada peringkat
- Domestication and Plant Genomes-2009Dokumen7 halamanDomestication and Plant Genomes-2009vahidvasharBelum ada peringkat
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- 2018 Air Pressure and Fronts PracticeDokumen10 halaman2018 Air Pressure and Fronts PracticeUlugbek SalohiddinovBelum ada peringkat
- DesertificationDokumen6 halamanDesertificationaminatariq0603Belum ada peringkat
- Crops of Northern PlainsDokumen6 halamanCrops of Northern Plainsitsankurz100% (1)
- Tugas B.inggris Kel.1Dokumen3 halamanTugas B.inggris Kel.1́AyuᴎBelum ada peringkat
- Presentation On Populas NigraDokumen41 halamanPresentation On Populas NigraMUHAMMAD ILYASBelum ada peringkat
- Deserts Around The WorldDokumen7 halamanDeserts Around The WorldAndreea V.A.F.Belum ada peringkat
- Climate Change Impact Assessment On Hydrology of IndianDokumen38 halamanClimate Change Impact Assessment On Hydrology of IndianarjunBelum ada peringkat
- Worksheet N º 6. Sixth Grade. Unit 7.: Colegio San SebastiánDokumen3 halamanWorksheet N º 6. Sixth Grade. Unit 7.: Colegio San Sebastiángenesis llancapaniBelum ada peringkat
- Chap 013Dokumen18 halamanChap 013Patrick FahrenbachBelum ada peringkat
- Uluru 2Dokumen12 halamanUluru 2api-250293174Belum ada peringkat
- Lect - 4Dokumen3 halamanLect - 4baigBelum ada peringkat
- Savanna Climate or Tropical Wet and Dry Climate or Sudan ClimateDokumen8 halamanSavanna Climate or Tropical Wet and Dry Climate or Sudan ClimateVikram DasBelum ada peringkat
- Geography 11 Week 1 PDFDokumen3 halamanGeography 11 Week 1 PDFNoah ZerihunBelum ada peringkat
- Flood RescueDokumen7 halamanFlood RescueDien NoelBelum ada peringkat
- Floating Ice: Assignment in Risk Readiness and Risk ReductionDokumen1 halamanFloating Ice: Assignment in Risk Readiness and Risk ReductionJoseph EstellosoBelum ada peringkat
- CC Presentation TAGUEMOUNTDokumen20 halamanCC Presentation TAGUEMOUNTRihab TAGUEMOUNTBelum ada peringkat
- Gilgamesh and Ishtar and The Death of EnkiduDokumen28 halamanGilgamesh and Ishtar and The Death of Enkidujackie0403Belum ada peringkat
- AlamadaDokumen2 halamanAlamadaAngeliza DavidBelum ada peringkat
- MidtemDokumen6 halamanMidtemkhahoaBelum ada peringkat
- Climatograph Biomes Worksheet 1Dokumen5 halamanClimatograph Biomes Worksheet 1Joanie GuthzeitBelum ada peringkat
- Detection of Heavy MetalsDokumen3 halamanDetection of Heavy MetalsAlphaRaj MekapoguBelum ada peringkat
- NCSCDokumen12 halamanNCSCkvians0% (1)
- Soal Conjunction Pilihan GandaDokumen2 halamanSoal Conjunction Pilihan GandaOke ImanBelum ada peringkat
- I. Read The Selection Below and Encircle The Letter of The Correct AnswerDokumen2 halamanI. Read The Selection Below and Encircle The Letter of The Correct AnswerGenerose Cantilan-Corvera Tabulo GarciaBelum ada peringkat
- Malawi Climate Change PresentationDokumen14 halamanMalawi Climate Change PresentationOxfam Youth and SchoolsBelum ada peringkat
- The Famine Game: Note To Pics: I Think, We Should Paste The Bookmarks in Front LaDokumen2 halamanThe Famine Game: Note To Pics: I Think, We Should Paste The Bookmarks in Front LaChong Ru YinBelum ada peringkat
- Final Report SRIDokumen21 halamanFinal Report SRIMinaketan RayBelum ada peringkat
- 388 19th Street Oakland, California 94612 DOA Phone LMUDA Phone 510.452.4529Dokumen11 halaman388 19th Street Oakland, California 94612 DOA Phone LMUDA Phone 510.452.4529OaklandCBDsBelum ada peringkat
- StrandedDokumen5 halamanStrandedapi-322303922Belum ada peringkat
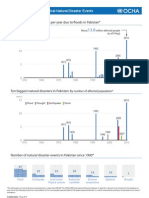
- Pakistan Floods-Historical Natural Disaster EventsDokumen1 halamanPakistan Floods-Historical Natural Disaster Eventsmarkleongoldber3215Belum ada peringkat