Anda mungkin juga menyukai
- 4 1 Operons Pos NegDokumen27 halaman4 1 Operons Pos NegAbdul RehemanBelum ada peringkat
- Regulation of Gene ExpressionDokumen53 halamanRegulation of Gene Expressionarman.laly82Belum ada peringkat
- Lecture 7 - Regulation of Gene Expression 05-22-12-1Dokumen25 halamanLecture 7 - Regulation of Gene Expression 05-22-12-1Lisa Brett HoffmanBelum ada peringkat
- David Yu Zhang and Georg Seelig - DNA-Based Fixed Gain Amplifiers and Linear Classifier CircuitsDokumen11 halamanDavid Yu Zhang and Georg Seelig - DNA-Based Fixed Gain Amplifiers and Linear Classifier CircuitsCogsmsBelum ada peringkat
- Gene Regulation in Prokaryotes: Presented by Dr. Nitin M. Kamble PHD Scholar BtyDokumen33 halamanGene Regulation in Prokaryotes: Presented by Dr. Nitin M. Kamble PHD Scholar BtyNitin KambleBelum ada peringkat
- TRP OperonDokumen80 halamanTRP OperonIsmi SitiBelum ada peringkat
- Gene Regulation Notes 1. Cybernetics (Steersman, Governor, Pilot or Rudder)Dokumen11 halamanGene Regulation Notes 1. Cybernetics (Steersman, Governor, Pilot or Rudder)emmyrubiscoBelum ada peringkat
- Gene RegulationDokumen13 halamanGene RegulationLeenakshi TyagiBelum ada peringkat
- 208 Euk Regulation of Gene Expression 2012Dokumen49 halaman208 Euk Regulation of Gene Expression 2012Ishtiaq Ahmed MawlaBelum ada peringkat
- BCH100 203 inter Gene regulation (2019-2) เอกสารDokumen54 halamanBCH100 203 inter Gene regulation (2019-2) เอกสารTsheden TshokeyBelum ada peringkat
- Footprinting DNA-Protein InteractionsDokumen18 halamanFootprinting DNA-Protein InteractionsSruthiBelum ada peringkat
- Prokaryotic Gene ExpressionDokumen44 halamanProkaryotic Gene ExpressionMelissaBelum ada peringkat
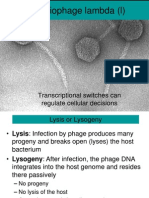
- Bacteriophage Lambda (L) : Transcriptional Switches Can Regulate Cellular DecisionsDokumen23 halamanBacteriophage Lambda (L) : Transcriptional Switches Can Regulate Cellular Decisionsmaniiiiiiii100% (1)
- Regulation of MetabolismDokumen66 halamanRegulation of MetabolismObinna OjeBelum ada peringkat
- Pakpoom Subsoontorn, Jongmin Kim and Erik Winfree - Bistability of An in Vitro Synthetic Autoregulatory SwitchDokumen21 halamanPakpoom Subsoontorn, Jongmin Kim and Erik Winfree - Bistability of An in Vitro Synthetic Autoregulatory SwitchCogsmsBelum ada peringkat
- Chapter 7 Control of Gene ExpressionDokumen80 halamanChapter 7 Control of Gene ExpressionLeonie Cruz Reyes100% (1)
- How To Study DNA: 1. Genetic Material 2. Expression ProductDokumen57 halamanHow To Study DNA: 1. Genetic Material 2. Expression ProductLaila Istighfaroh Ella EllEllBelum ada peringkat
- Lodish6e ch07Dokumen8 halamanLodish6e ch07nBelum ada peringkat
- Molecular Biology (Genes and Mutations)Dokumen61 halamanMolecular Biology (Genes and Mutations)Justin PRUDENCIOBelum ada peringkat
- Test 3 Study Guide BSC 315Dokumen2 halamanTest 3 Study Guide BSC 315MelanieBelum ada peringkat
- 12 Regulasi Ekspresi Gen Pada EukariotDokumen27 halaman12 Regulasi Ekspresi Gen Pada EukariotvinaBelum ada peringkat
- Gateway Technology: M.Phil. Biotechnology CBM, University of SwatDokumen9 halamanGateway Technology: M.Phil. Biotechnology CBM, University of SwatGul AfshaBelum ada peringkat
- Gene Regulation in BacteriaDokumen51 halamanGene Regulation in BacteriaJoyghoshtotradeBelum ada peringkat
- Molecular Microbiology MAP Prep FinalDokumen41 halamanMolecular Microbiology MAP Prep FinalAdi SassonBelum ada peringkat
- Part 4 Case Study - Industrial EnzymesDokumen33 halamanPart 4 Case Study - Industrial EnzymesyahmedpersBelum ada peringkat
- Gene Regulation in Prokaryotes-1Dokumen17 halamanGene Regulation in Prokaryotes-1Fasiha Mushadi100% (1)
- Chapt07 Image v1.8Dokumen82 halamanChapt07 Image v1.8brandon LUBelum ada peringkat
- Sect 9 Regulation Eu Kary OtesDokumen9 halamanSect 9 Regulation Eu Kary OtesNika AndrianiBelum ada peringkat
- Enzyme KineticsDokumen10 halamanEnzyme Kineticskedirwoliy100% (1)
- Real Time PCR: Using Sybr GreenDokumen78 halamanReal Time PCR: Using Sybr GreenMazin ElaminBelum ada peringkat
- F - Gateway Cloning TechnologyDokumen17 halamanF - Gateway Cloning TechnologyCarlos Alfredo Carmona GascaBelum ada peringkat
- Introduction To Genetic Analysis: Regulation of Gene Expression in Bacteria and Their VirusesDokumen46 halamanIntroduction To Genetic Analysis: Regulation of Gene Expression in Bacteria and Their VirusesMuneeb ArshadBelum ada peringkat
- Glycolysis and TCA Cycle ActivityDokumen6 halamanGlycolysis and TCA Cycle ActivityYannis ZoldenbergBelum ada peringkat
- Gene Expression in Prokaryotes.Dokumen23 halamanGene Expression in Prokaryotes.M.PRASAD NAIDU100% (1)
- Signal Transduction Pathways: Pratt & Cornely, Chapter 10Dokumen23 halamanSignal Transduction Pathways: Pratt & Cornely, Chapter 10Brother GeorgeBelum ada peringkat
- Gene Regulation in Prokaryotes 0Dokumen67 halamanGene Regulation in Prokaryotes 0Angel TaylorBelum ada peringkat
- Information Readout: TranscriptionDokumen53 halamanInformation Readout: TranscriptiongraceBelum ada peringkat
- Transcription Prokaryotes 2012-cDokumen27 halamanTranscription Prokaryotes 2012-cAnupama PatiBelum ada peringkat
- Promoters and Plasmids PDFDokumen4 halamanPromoters and Plasmids PDFAndika MardiantoBelum ada peringkat
- Directed Mutagenesis and Protein EngineeringDokumen47 halamanDirected Mutagenesis and Protein EngineeringNurulBelum ada peringkat
- Regulation of Gene ExpressionDokumen5 halamanRegulation of Gene ExpressionFathimaBelum ada peringkat
- Gene Regulation in ProkaryotesDokumen48 halamanGene Regulation in ProkaryotesJT3Belum ada peringkat
- 09 Regulation of Gene ExpressionDokumen42 halaman09 Regulation of Gene ExpressionFirst Anonym877Belum ada peringkat
- Biomol BiomedikFera Ibrahim08032018Dokumen95 halamanBiomol BiomedikFera Ibrahim08032018PMIB Matrikulasi FKUI 2018/2019Belum ada peringkat
- Scotty Merrell Department of Microbiology and Immunology B4140 Dmerrell@usuhs - MilDokumen70 halamanScotty Merrell Department of Microbiology and Immunology B4140 Dmerrell@usuhs - MilJT3Belum ada peringkat
- Chuong 4 TeamDokumen91 halamanChuong 4 Teamvuhust124Belum ada peringkat
- Transcription FactorsDokumen25 halamanTranscription FactorsPriya.RBelum ada peringkat
- Decker 1Dokumen9 halamanDecker 1Katarina Kaca GacevicBelum ada peringkat
- Regulation of Gene ExpressionDokumen34 halamanRegulation of Gene ExpressionJay Fickle100% (1)
- Regulation of Gene Expression-1Dokumen30 halamanRegulation of Gene Expression-1mus zaharaBelum ada peringkat
- Regulación Por AtenuaciónDokumen5 halamanRegulación Por AtenuaciónKaryme Aylín Martinez OliverosBelum ada peringkat
- 11TrpAMO LicyayoDokumen8 halaman11TrpAMO LicyayoMohamidin MamalapatBelum ada peringkat
- RNA TranscriptionDokumen37 halamanRNA Transcriptionarman.laly82Belum ada peringkat
- Molecular Cell Biology Lodish 6th Edition Test Bank DownloadDokumen9 halamanMolecular Cell Biology Lodish 6th Edition Test Bank Downloadjohndanielswrydmzokge100% (26)
- Gene RegulationsDokumen11 halamanGene Regulationsmahadi.infsduBelum ada peringkat
- CBT 2 Paper PDFDokumen140 halamanCBT 2 Paper PDFkarthikBelum ada peringkat
- Biochem Lec20Dokumen4 halamanBiochem Lec20Louis FortunatoBelum ada peringkat
- Signal Transduction: Key TopicsDokumen39 halamanSignal Transduction: Key TopicsBellony Sanders100% (1)
- Issues Related To Patenting of Biological InventionDokumen19 halamanIssues Related To Patenting of Biological InventionPavithra ShanmugamBelum ada peringkat
- Biotechnology Patenting in India and Related IssuesDokumen18 halamanBiotechnology Patenting in India and Related IssuesPankaj RathiBelum ada peringkat
- Application of Molecular Biology Knowledge in Developing New DrugsDokumen4 halamanApplication of Molecular Biology Knowledge in Developing New DrugsPavithra ShanmugamBelum ada peringkat
- The Neem Patent IssueDokumen4 halamanThe Neem Patent IssuePavithra ShanmugamBelum ada peringkat
- Pancreatic Cancer Therapy Using Engineered Mesenchymal Stem CellsDokumen12 halamanPancreatic Cancer Therapy Using Engineered Mesenchymal Stem CellsPavithra ShanmugamBelum ada peringkat
- Genetically Encoded Fluorescent Redox ProbesDokumen16 halamanGenetically Encoded Fluorescent Redox ProbesPavithra ShanmugamBelum ada peringkat
- Genetically Encoded Fluorescent Redox ProbesDokumen16 halamanGenetically Encoded Fluorescent Redox ProbesPavithra ShanmugamBelum ada peringkat
- Genetically Encoded Fluorescent Redox ProbesDokumen16 halamanGenetically Encoded Fluorescent Redox ProbesPavithra ShanmugamBelum ada peringkat
- Fighting Cancer With Vaccinia VirusDokumen21 halamanFighting Cancer With Vaccinia VirusPavithra ShanmugamBelum ada peringkat
- Ratner - Nanotechnology - A Gentle Introduction To The Next Big IdeaDokumen153 halamanRatner - Nanotechnology - A Gentle Introduction To The Next Big IdeaKo NwayBelum ada peringkat
- LC 67002000 B Hr-Jumbo enDokumen2 halamanLC 67002000 B Hr-Jumbo enJulio OrtegaBelum ada peringkat
- Lesson Tasks: Lesson Plan: Addition and Subtraction To 20Dokumen2 halamanLesson Tasks: Lesson Plan: Addition and Subtraction To 20Марина СтанојевићBelum ada peringkat
- VDA Volume Assessment of Quality Management Methods Guideline 1st Edition November 2017 Online-DocumentDokumen36 halamanVDA Volume Assessment of Quality Management Methods Guideline 1st Edition November 2017 Online-DocumentR JBelum ada peringkat
- P&CDokumen18 halamanP&Cmailrgn2176Belum ada peringkat
- Physical Characteristics of SoilDokumen26 halamanPhysical Characteristics of SoillfpachecoBelum ada peringkat
- Solutions - HW 3, 4Dokumen5 halamanSolutions - HW 3, 4batuhany90Belum ada peringkat
- ARMY - Chapter Eight Explosive BreachingDokumen25 halamanARMY - Chapter Eight Explosive Breachingrsreeth100% (1)
- American University of Beirut PSPA 210: Intro. To Political ThoughtDokumen4 halamanAmerican University of Beirut PSPA 210: Intro. To Political Thoughtcharles murrBelum ada peringkat
- Section ADokumen7 halamanSection AZeeshan HaiderBelum ada peringkat
- Business Mathematics (Matrix)Dokumen3 halamanBusiness Mathematics (Matrix)MD HABIBBelum ada peringkat
- Attitude of Tribal and Non Tribal Students Towards ModernizationDokumen9 halamanAttitude of Tribal and Non Tribal Students Towards ModernizationAnonymous CwJeBCAXpBelum ada peringkat
- 2019 Auto Truck Key Blank Reference PDFDokumen188 halaman2019 Auto Truck Key Blank Reference PDFAlbert RodaBelum ada peringkat
- Leonard Nadler' ModelDokumen3 halamanLeonard Nadler' ModelPiet Gabz67% (3)
- Part TOEFLDokumen7 halamanPart TOEFLFrisca Rahma DwinantiBelum ada peringkat
- Army Public School No.1 Jabalpur Practical List - Computer Science Class - XIIDokumen4 halamanArmy Public School No.1 Jabalpur Practical List - Computer Science Class - XIIAdityaBelum ada peringkat
- Procedure Manual - IMS: Locomotive Workshop, Northern Railway, LucknowDokumen8 halamanProcedure Manual - IMS: Locomotive Workshop, Northern Railway, LucknowMarjorie Dulay Dumol80% (5)
- TOPIC 2 - Fans, Blowers and Air CompressorDokumen69 halamanTOPIC 2 - Fans, Blowers and Air CompressorCllyan ReyesBelum ada peringkat
- Early Childhood Education and CareDokumen53 halamanEarly Childhood Education and CareBianca ALbuquerqueBelum ada peringkat
- Hasan Bin Ekram: Career ObjectiveDokumen3 halamanHasan Bin Ekram: Career ObjectiveHasan SarikBelum ada peringkat
- Angles, Bearings and AzimuthDokumen10 halamanAngles, Bearings and AzimuthMarc Dared CagaoanBelum ada peringkat
- Open Book Online: Syllabus & Pattern Class - XiDokumen1 halamanOpen Book Online: Syllabus & Pattern Class - XiaadityaBelum ada peringkat
- Dosificación Gac007-008 Sem2Dokumen2 halamanDosificación Gac007-008 Sem2Ohm EgaBelum ada peringkat
- Chapter5A TorqueDokumen32 halamanChapter5A TorqueShuq Faqat al-FansuriBelum ada peringkat
- Cooperative LinuxDokumen39 halamanCooperative Linuxrajesh_124Belum ada peringkat
- What Role Does Imagination Play in Producing Knowledge About The WorldDokumen1 halamanWhat Role Does Imagination Play in Producing Knowledge About The WorldNathanael Samuel KuruvillaBelum ada peringkat
- STIHL TS410, TS420 Spare PartsDokumen11 halamanSTIHL TS410, TS420 Spare PartsMarinko PetrovićBelum ada peringkat
- Paper 11-ICOSubmittedDokumen10 halamanPaper 11-ICOSubmittedNhat Tan MaiBelum ada peringkat
- JurnalDokumen12 halamanJurnalSandy Ronny PurbaBelum ada peringkat
- RF Based Dual Mode RobotDokumen17 halamanRF Based Dual Mode Robotshuhaibasharaf100% (2)
- Archetypal Approach To Macbeth by William ShakespeareDokumen9 halamanArchetypal Approach To Macbeth by William ShakespeareLenka Koutná100% (1)