Anda mungkin juga menyukai
- Watson and Raynor 1920Dokumen9 halamanWatson and Raynor 1920ShaneLee10267% (3)
- The Effects of Training On Fatigue and Twitch Potentiation in Human Skeletal MuscleDokumen9 halamanThe Effects of Training On Fatigue and Twitch Potentiation in Human Skeletal MuscleCharisBelum ada peringkat
- Muscle and Exercise PhysiologyDari EverandMuscle and Exercise PhysiologyJerzy A. ZoladzPenilaian: 3.5 dari 5 bintang3.5/5 (3)
- Muscle Nerve,: 1997 John Wiley & Sons, Inc. 20, 336-346, 1997Dokumen11 halamanMuscle Nerve,: 1997 John Wiley & Sons, Inc. 20, 336-346, 1997sunshinefreshBelum ada peringkat
- Anaerobic MetabolismDokumen10 halamanAnaerobic MetabolismPetros GrigorakisBelum ada peringkat
- In Vivo Reduction in ATP Cost of Contraction Is Not RelatedDokumen11 halamanIn Vivo Reduction in ATP Cost of Contraction Is Not RelatedCharisBelum ada peringkat
- Mitchell Et Al. (2009) Reflex Mechanisms Lack of ContributionDokumen16 halamanMitchell Et Al. (2009) Reflex Mechanisms Lack of ContributionRafael RodriguesBelum ada peringkat
- Conley 1998Dokumen11 halamanConley 1998M Al-FathBelum ada peringkat
- I P P I I R E P: Schemic Reconditioning and Lacebo Ntervention Mproves Esistance Xercise ErformanceDokumen8 halamanI P P I I R E P: Schemic Reconditioning and Lacebo Ntervention Mproves Esistance Xercise ErformanceBatta ZahranBelum ada peringkat
- Effects of Creatine Supplementation On The Energy Cost of Muscle ContractionDokumen9 halamanEffects of Creatine Supplementation On The Energy Cost of Muscle ContractionCharisBelum ada peringkat
- Paper BioenergéticaDokumen10 halamanPaper BioenergéticaReycel Lapierre PobleteBelum ada peringkat
- Bertil Sjodin Et - Al, 1982 - Changes in Onset of Blood Lactate Accumulation (OBLA) and Muscle Enzymes After Training at OBLADokumen13 halamanBertil Sjodin Et - Al, 1982 - Changes in Onset of Blood Lactate Accumulation (OBLA) and Muscle Enzymes After Training at OBLAok okBelum ada peringkat
- Effect of Muscle Oxygenation During Resistance Exercise On Anabolic Hormone ResponseDokumen6 halamanEffect of Muscle Oxygenation During Resistance Exercise On Anabolic Hormone ResponseAle CostaBelum ada peringkat
- Bogdanis-1996 Contribution of Phosphocreatine and Aerobic Metabolism To Energy Supply During RsaDokumen9 halamanBogdanis-1996 Contribution of Phosphocreatine and Aerobic Metabolism To Energy Supply During Rsapreparador fisicoBelum ada peringkat
- Skeletal Troponin I As A Marker of Exercise-Induced Muscle DamageDokumen7 halamanSkeletal Troponin I As A Marker of Exercise-Induced Muscle DamagedbcooperdkBelum ada peringkat
- NG 2007Dokumen8 halamanNG 2007Nacho MicheloniBelum ada peringkat
- Locomotor Muscle Fatigue Modifies Central Motor Drive in Healthy Humans and Imposes A Limitation To Exercise PerformanceDokumen13 halamanLocomotor Muscle Fatigue Modifies Central Motor Drive in Healthy Humans and Imposes A Limitation To Exercise PerformanceRei ArturBelum ada peringkat
- Jurnal Contrax RelaxDokumen8 halamanJurnal Contrax Relaxfi.afifah NurBelum ada peringkat
- Programme of Master Education (Biology) : Lecturer: DR Wong Chee FahDokumen19 halamanProgramme of Master Education (Biology) : Lecturer: DR Wong Chee Fahsoonzi04Belum ada peringkat
- Müller, 2016Dokumen8 halamanMüller, 2016Pablo fernandezBelum ada peringkat
- Applied Physiology: J. M. Pivarnik, M. P. Goetting, and L. C. Senay, JRDokumen7 halamanApplied Physiology: J. M. Pivarnik, M. P. Goetting, and L. C. Senay, JRWAI SAM LIMBelum ada peringkat
- Skeletal Muscle Signaling Responses To Resistance Exercise of The Elbow Extensors Are Not Compromised by A Preceding Bout of Aerobic ExerciseDokumen35 halamanSkeletal Muscle Signaling Responses To Resistance Exercise of The Elbow Extensors Are Not Compromised by A Preceding Bout of Aerobic Exerciseleal thiagoBelum ada peringkat
- Tugas Kelompok Ke 2Dokumen1 halamanTugas Kelompok Ke 2Khykhy Rezskhy MuliyanaBelum ada peringkat
- High Level of Skeletal Muscle Carnosine Contributes To The Latter Half of Exercise Performance During 30-s Maximal Cycle Ergometer SprintingDokumen7 halamanHigh Level of Skeletal Muscle Carnosine Contributes To The Latter Half of Exercise Performance During 30-s Maximal Cycle Ergometer Sprintinglika-chanBelum ada peringkat
- Dosagem Otima Musculo PDFDokumen10 halamanDosagem Otima Musculo PDFJanaina PanizzaBelum ada peringkat
- Effect of Training On Muscle Metabolism During Treadmill SprintingDokumen7 halamanEffect of Training On Muscle Metabolism During Treadmill SprintingBRIAN JUMA NYONGESABelum ada peringkat
- Pullover 2018Dokumen6 halamanPullover 2018Lina M GarciaBelum ada peringkat
- Caffeine Ingestion Reverses The Circadian Rhythm Effects On Neuromuscular Performance in Highly Resistance-Trained Men - Mora-Rodríguez 2012Dokumen9 halamanCaffeine Ingestion Reverses The Circadian Rhythm Effects On Neuromuscular Performance in Highly Resistance-Trained Men - Mora-Rodríguez 2012Yo Vivo Fit Pablo y KarlaBelum ada peringkat
- Bioquimica 2Dokumen7 halamanBioquimica 2FerroBemBelum ada peringkat
- The Acute Muscle Swelling Effects of Blood Flow RestrictionDokumen11 halamanThe Acute Muscle Swelling Effects of Blood Flow RestrictionFabiano LacerdaBelum ada peringkat
- Trump Et Al 2001 An 1h Mrs Evaluation of The Phosphocreatine Creatine Pool (TCR) in Human MuscleDokumen8 halamanTrump Et Al 2001 An 1h Mrs Evaluation of The Phosphocreatine Creatine Pool (TCR) in Human MuscleКолевска МиленаBelum ada peringkat
- Articulo #30Dokumen8 halamanArticulo #30LAURA CAMILA CUBILLOS VILLAMILBelum ada peringkat
- JSCR 19 2 421-426 2005 Stretching StrengthDokumen6 halamanJSCR 19 2 421-426 2005 Stretching StrengthFernanda KerlenBelum ada peringkat
- Effect of An Acute Period of Resistance Exercise On Excess Post-Exercise Oxygen ConsumptionDokumen7 halamanEffect of An Acute Period of Resistance Exercise On Excess Post-Exercise Oxygen ConsumptionaxkielyBelum ada peringkat
- Gaitanos 1993 PDFDokumen8 halamanGaitanos 1993 PDFLeonardo AlvesBelum ada peringkat
- Ajpendo 00387 2004Dokumen7 halamanAjpendo 00387 2004Diego Duarte CavalcanteBelum ada peringkat
- Stretching PhisiologyDokumen6 halamanStretching PhisiologyHnk Bjelopoljac Škola NogometaBelum ada peringkat
- Ankle Cryotherapy Facilitates SoleusDokumen6 halamanAnkle Cryotherapy Facilitates SoleusRamiro Avendaño RebolledoBelum ada peringkat
- Reaction Paper 1Dokumen3 halamanReaction Paper 1Erna GampalBelum ada peringkat
- Adam 05Dokumen14 halamanAdam 05MartinVertolazBelum ada peringkat
- 2002 The Yo-Yo Intermittent Recovery Test Physiological Response, Reliability, and ValidityDokumen9 halaman2002 The Yo-Yo Intermittent Recovery Test Physiological Response, Reliability, and ValidityFrancisco Antonó Castro WeithBelum ada peringkat
- JMD 19083Dokumen3 halamanJMD 19083anton MDBelum ada peringkat
- Proprioceptive Neuromuscular Facilitation (PNF) Stretching Techniques Hold-Relax HR ALLDokumen8 halamanProprioceptive Neuromuscular Facilitation (PNF) Stretching Techniques Hold-Relax HR ALLKiné Therapeut-manuelle Masseur GabiBelum ada peringkat
- Contribution of Phosphocreatine and Aerobic Metabolism To Energy Supply During Repeated Sprint ExerciseDokumen10 halamanContribution of Phosphocreatine and Aerobic Metabolism To Energy Supply During Repeated Sprint ExerciseEduardo VieyraBelum ada peringkat
- T A R P O K E: HE Cute Esponse of Ractical Cclusion in THE NEE XtensorsDokumen4 halamanT A R P O K E: HE Cute Esponse of Ractical Cclusion in THE NEE XtensorsJuanfran Montesinos GarciaBelum ada peringkat
- Wavelet Analysis of Rearfoot Loading of Patients After Medial Patellofemoral Ligament MPFL ReconstructionDokumen6 halamanWavelet Analysis of Rearfoot Loading of Patients After Medial Patellofemoral Ligament MPFL ReconstructionceydaalyazhotamisBelum ada peringkat
- Abnormal Mitochondrial Function and Muscle Wasting, But Normal Contractile Efficiency, in Haemodialysed Patients Studied Non-Invasively in VivoDokumen8 halamanAbnormal Mitochondrial Function and Muscle Wasting, But Normal Contractile Efficiency, in Haemodialysed Patients Studied Non-Invasively in VivoDaniel Fernando Mendez CarbajalBelum ada peringkat
- Effects of Caffeine On Session Ratings of Perceived ExertionDokumen7 halamanEffects of Caffeine On Session Ratings of Perceived ExertionJackson PierceBelum ada peringkat
- Objective Evaluation of A Splint Designed To Reduce H YpertonicityDokumen6 halamanObjective Evaluation of A Splint Designed To Reduce H YpertonicityNoor efBelum ada peringkat
- Stretching On VO2 MaxDokumen12 halamanStretching On VO2 MaxEli J. TorresBelum ada peringkat
- Acute Effect of Two Aerobic Exercise ModesDokumen5 halamanAcute Effect of Two Aerobic Exercise Modeswander_dukaBelum ada peringkat
- TMP F204Dokumen2 halamanTMP F204FrontiersBelum ada peringkat
- 24th Annual ECSS Congress Prague/Czech Republic, July 3-6 2019Dokumen1 halaman24th Annual ECSS Congress Prague/Czech Republic, July 3-6 2019Karini Borges Dos SantosBelum ada peringkat
- Naclerio2019 Article EffectivenessOfCombiningMicrocDokumen13 halamanNaclerio2019 Article EffectivenessOfCombiningMicrocAna SandovalBelum ada peringkat
- Electromyographic Activation of Pectoralis Major and Triceps BrachiiDokumen11 halamanElectromyographic Activation of Pectoralis Major and Triceps BrachiiLina M GarciaBelum ada peringkat
- Parasympathetic Reactivation After Repeated Sprint Exercise: Martin Buchheit, Paul B. Laursen, and Saı D AhmaidiDokumen9 halamanParasympathetic Reactivation After Repeated Sprint Exercise: Martin Buchheit, Paul B. Laursen, and Saı D AhmaidiVeronica JanethBelum ada peringkat
- ARTICLE-Effects of A Wheelchair Ergometer Training Programme On SpinalDokumen6 halamanARTICLE-Effects of A Wheelchair Ergometer Training Programme On SpinalavalosheBelum ada peringkat
- Absctrac Congreso NSCADokumen144 halamanAbsctrac Congreso NSCAMelina LabroneBelum ada peringkat
- Wang Et Al. (1999)Dokumen5 halamanWang Et Al. (1999)Jay CHANBelum ada peringkat
- Better BalanceDokumen13 halamanBetter BalanceMiriam NovoBelum ada peringkat
- Biochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967Dari EverandBiochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967D. RichterBelum ada peringkat
- Network ListDokumen880 halamanNetwork ListRobbin BajpaiBelum ada peringkat
- Intensified Testing For Attention-Deficit Hyperactivity Disorder (ADHD) in Girls Should Reduce Depression and Smoking in Adult Females ADokumen4 halamanIntensified Testing For Attention-Deficit Hyperactivity Disorder (ADHD) in Girls Should Reduce Depression and Smoking in Adult Females ARafael MartinsBelum ada peringkat
- GoniosDokumen55 halamanGoniosdrquan100% (1)
- 2017 - Resume BaseDokumen3 halaman2017 - Resume Baseapi-341527743Belum ada peringkat
- Case Report Two Blue Chromhidrosis Patients in Emergency RoomDokumen2 halamanCase Report Two Blue Chromhidrosis Patients in Emergency RoomInternational Journal of Innovative Science and Research TechnologyBelum ada peringkat
- JanitaDokumen2 halamanJanitaapi-391376321Belum ada peringkat
- Developmental Dysplasia of The HipDokumen5 halamanDevelopmental Dysplasia of The HipMavra zBelum ada peringkat
- Complications of Tonsillectomy, Adenoidectomy and Adenotonsillectomy in Chittagong Medical College HospitalDokumen4 halamanComplications of Tonsillectomy, Adenoidectomy and Adenotonsillectomy in Chittagong Medical College Hospitalnaufal12345Belum ada peringkat
- Glossary EyeDokumen228 halamanGlossary EyeCarolus AugustusBelum ada peringkat
- Vaginal Birth After Previous Caeserian SectionDokumen18 halamanVaginal Birth After Previous Caeserian SectionSky JuiceBelum ada peringkat
- Pediatric Medication Math Review Jan 2011Dokumen7 halamanPediatric Medication Math Review Jan 2011Tyrone Kent HalogBelum ada peringkat
- Current Diagnosis Treatment Pediatrics 24Th Edition Edition William Hay Full ChapterDokumen67 halamanCurrent Diagnosis Treatment Pediatrics 24Th Edition Edition William Hay Full Chapterbill.groff554100% (4)
- Tapentadol ParacetamolDokumen2 halamanTapentadol ParacetamolMohammad Shahbaz Alam100% (1)
- Ped 10Dokumen17 halamanPed 10Tufail AhmadBelum ada peringkat
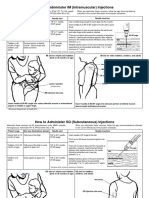
- Im SQ Admin PDFDokumen2 halamanIm SQ Admin PDFAlvin JjBelum ada peringkat
- Mta Vs Formocresol PulpotomyDokumen4 halamanMta Vs Formocresol PulpotomySitapriya SubrahmanyamBelum ada peringkat
- Hirschprung Associated EnterocolitisDokumen9 halamanHirschprung Associated Enterocolitiskeyla_shineeeBelum ada peringkat
- Huntsville Businesses That Received More Than $150,000 From The Paycheck Protection ProgramDokumen14 halamanHuntsville Businesses That Received More Than $150,000 From The Paycheck Protection ProgramJonathan GrassBelum ada peringkat
- Aniapravan 2020Dokumen2 halamanAniapravan 2020Marcos HernandezBelum ada peringkat
- Oily Skin 1Dokumen7 halamanOily Skin 1GligorpaulaBelum ada peringkat
- Unit 4 Management of Open Pulp in Deciduous TeethDokumen11 halamanUnit 4 Management of Open Pulp in Deciduous Teethisti DaristiviaBelum ada peringkat
- NMB MyDokumen91 halamanNMB MyGirish Bk B KBelum ada peringkat
- Atlas of Osteopathic TechniquesDokumen81 halamanAtlas of Osteopathic TechniquesDanieleBelum ada peringkat
- Atypical EclampsiaDokumen3 halamanAtypical EclampsiadianaristinugraheniBelum ada peringkat
- Acute Nephrology For The Critical Care Physician 2015th EditionDokumen280 halamanAcute Nephrology For The Critical Care Physician 2015th EditionHiago Sousa100% (1)
- Jurnal ReadingDokumen22 halamanJurnal Readingarismaputra20Belum ada peringkat
- Spek Ven Avea Full ClioDokumen3 halamanSpek Ven Avea Full ClioRinawatiBelum ada peringkat
- B5W1L9.Peripheral Neuropathy - Lecture Notes 12Dokumen4 halamanB5W1L9.Peripheral Neuropathy - Lecture Notes 12mihalcea alinBelum ada peringkat
- 1 Childhood Nephrotic Syndrome - Diagnosis and ManagementDokumen52 halaman1 Childhood Nephrotic Syndrome - Diagnosis and ManagementThana Balan100% (1)