Anda mungkin juga menyukai
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- A Study of The Electrochemical Oxidation and Polymerisation of 1,3 Dihydroxybenzene and 3-Hydroxybenzyl Alcohol in Acidic, Basic and Neutral Aqueous SolutionsDokumen11 halamanA Study of The Electrochemical Oxidation and Polymerisation of 1,3 Dihydroxybenzene and 3-Hydroxybenzyl Alcohol in Acidic, Basic and Neutral Aqueous SolutionsEdward PittsBelum ada peringkat
- 8 and 15°C.: Stability ProgramsDokumen1 halaman8 and 15°C.: Stability ProgramsEdward PittsBelum ada peringkat
- Electrochemical Oxidation of Paracetamol Mediated by Nanoparticles Bismuth Oxide Modified Glassy Carbon ElectrodeDokumen10 halamanElectrochemical Oxidation of Paracetamol Mediated by Nanoparticles Bismuth Oxide Modified Glassy Carbon ElectrodeEdward PittsBelum ada peringkat
- 5990 7890en PDFDokumen4 halaman5990 7890en PDFEdward PittsBelum ada peringkat
- Chemical Space Travel: Ruud Van Deursen and Jean-Louis ReymondDokumen5 halamanChemical Space Travel: Ruud Van Deursen and Jean-Louis ReymondEdward PittsBelum ada peringkat
- Stats ch10 PDFDokumen14 halamanStats ch10 PDFEdward PittsBelum ada peringkat
- Vocabuilder ThesisDokumen32 halamanVocabuilder ThesisEdward PittsBelum ada peringkat
- Differential Calculus: 61 Differentiation of ProcessesDokumen67 halamanDifferential Calculus: 61 Differentiation of ProcessesEdward PittsBelum ada peringkat
- 170 - 058 - Conservation of Medicinal Plants in BangladeshDokumen22 halaman170 - 058 - Conservation of Medicinal Plants in BangladeshEdward PittsBelum ada peringkat
- Chemistry and Biotechnology: A Productive Union Meets New ChallengesDokumen3 halamanChemistry and Biotechnology: A Productive Union Meets New ChallengesEdward PittsBelum ada peringkat
- Spectroscopy NMR, Ir, MS, Uv-Vis: Main Points of The ChapterDokumen22 halamanSpectroscopy NMR, Ir, MS, Uv-Vis: Main Points of The ChapterEdward PittsBelum ada peringkat
- 03 GenX RaysDokumen14 halaman03 GenX RaysEdward PittsBelum ada peringkat
- 125Dokumen3 halaman125Edward Pitts27% (11)
- Search For Salt Colors: Ed BowmanDokumen4 halamanSearch For Salt Colors: Ed BowmanEdward Pitts100% (2)
- Long Distance Electron Transfer in Cytochrome C Oxidase Immobilised On Electrodes. A Surface Enhanced Resonance Raman Spectroscopic StudyDokumen8 halamanLong Distance Electron Transfer in Cytochrome C Oxidase Immobilised On Electrodes. A Surface Enhanced Resonance Raman Spectroscopic StudyEdward PittsBelum ada peringkat
- Spectral Properties, Generation Order Parameters, and Luminosities For Spin-Powered X-Ray PulsarsDokumen7 halamanSpectral Properties, Generation Order Parameters, and Luminosities For Spin-Powered X-Ray PulsarsEdward PittsBelum ada peringkat
- Ionization Masses: Molecular Ion PeakDokumen24 halamanIonization Masses: Molecular Ion PeakEdward PittsBelum ada peringkat
- Mass SpectrosDokumen47 halamanMass SpectrosEdward PittsBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5795)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1091)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Unsur Unsur Pokok Pelaksana Pembangunan Suatu ProyekDokumen15 halamanUnsur Unsur Pokok Pelaksana Pembangunan Suatu ProyekAjie'BondBelum ada peringkat
- Folic Acid and FolatesDokumen443 halamanFolic Acid and FolatesSuyog patilBelum ada peringkat
- La Vie en Rose'': Biosynthesis, Sources, and Applications of Betalain PigmentsDokumen16 halamanLa Vie en Rose'': Biosynthesis, Sources, and Applications of Betalain PigmentsNemes BiancaBelum ada peringkat
- Chemistry Fy23 Titles UkDokumen10 halamanChemistry Fy23 Titles Uknoura aldayanBelum ada peringkat
- Cell Transport: Lesson QuizDokumen3 halamanCell Transport: Lesson QuizAlberto ContechaBelum ada peringkat
- Mitochondria Genome OrganisationDokumen7 halamanMitochondria Genome OrganisationSushma KannapiranBelum ada peringkat
- Biologia Celular y MolecularDokumen24 halamanBiologia Celular y Molecularjohntc100% (1)
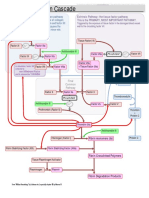
- Coagulation CascadeDokumen5 halamanCoagulation Cascadepieterinpretoria391100% (1)
- Medicina Mitocondrial Vol 1Dokumen485 halamanMedicina Mitocondrial Vol 1ArianPedrozaBelum ada peringkat
- EXPERIMENT 5 - Cloning Clone JET PCR CloningDokumen7 halamanEXPERIMENT 5 - Cloning Clone JET PCR CloningMirza IrfanBelum ada peringkat
- Natural History of HIV Infection by DR A K Gupta, Addl. Project Director, DSACSDokumen29 halamanNatural History of HIV Infection by DR A K Gupta, Addl. Project Director, DSACSdr.a.k.gupta6924100% (3)
- Gene Cloning & Cloning Vectors: DR Ravi Kant Agrawal, MVSC, PHDDokumen75 halamanGene Cloning & Cloning Vectors: DR Ravi Kant Agrawal, MVSC, PHDvivekBelum ada peringkat
- Yasko Methylation Pathway: Methionine Cycle Folate CycleDokumen6 halamanYasko Methylation Pathway: Methionine Cycle Folate CycleEric Ehle100% (1)
- HW 1 PDFDokumen3 halamanHW 1 PDFsuudfiinBelum ada peringkat
- Meiosis: Chapter 19 Pages 651 - 663Dokumen20 halamanMeiosis: Chapter 19 Pages 651 - 663María SalazarBelum ada peringkat
- Genetics (Biol 105) Linkage and Recombination Exercises: Progeny Phenotype Progeny NumberDokumen4 halamanGenetics (Biol 105) Linkage and Recombination Exercises: Progeny Phenotype Progeny NumberAngelica GeneraleBelum ada peringkat
- Obeppar Medical SlidesDokumen52 halamanObeppar Medical SlidesNimesh ModiBelum ada peringkat
- Experiment LipidsDokumen5 halamanExperiment LipidsSHAFIKANOR3661Belum ada peringkat
- Hexuronato Cita deDokumen13 halamanHexuronato Cita deDiegoBelum ada peringkat
- DPP 1 Cell BiologyDokumen2 halamanDPP 1 Cell Biology1149 Vanshika BBelum ada peringkat
- Asn of Genetics by Umair 26Dokumen7 halamanAsn of Genetics by Umair 26sdfksssglksfdfgsBelum ada peringkat
- M.sc. Applied MicrobiologyDokumen18 halamanM.sc. Applied Microbiologyprism1702Belum ada peringkat
- 10 3389@fimmu 2020 01100 PDFDokumen22 halaman10 3389@fimmu 2020 01100 PDFvalinaBelum ada peringkat
- Biologc3ada La Vida en La Tierra Con Fisiologc3ada 9c2ba Edicic3b3nDokumen2 halamanBiologc3ada La Vida en La Tierra Con Fisiologc3ada 9c2ba Edicic3b3nLaura Teresa PinzónBelum ada peringkat
- Stages of MeiosisDokumen4 halamanStages of MeiosisNicole CabrigasBelum ada peringkat
- I - in Vivo - I - and - I - in PDFDokumen4 halamanI - in Vivo - I - and - I - in PDFNatasha VaniaBelum ada peringkat
- Biotechnology SyllabusDokumen19 halamanBiotechnology SyllabuslmfaoBelum ada peringkat
- Zinc Overview PDFDokumen11 halamanZinc Overview PDFLê Nguyễn Hoàng Anh100% (1)
- Biophysics & Molecular Biology by Pranav KumarDokumen200 halamanBiophysics & Molecular Biology by Pranav KumarWajid Riyaz85% (54)
- Genomic Dna Isolation From Human Whole Blood Samples by Non Enzymatic Salting Out MethodDokumen2 halamanGenomic Dna Isolation From Human Whole Blood Samples by Non Enzymatic Salting Out MethodamandbhaskarBelum ada peringkat