Anda mungkin juga menyukai
- Eometry and RT: Ymmetry Alance CaleDokumen12 halamanEometry and RT: Ymmetry Alance CalemashhoodBelum ada peringkat
- Eometry and RT: Ymmetry Alance CaleDokumen12 halamanEometry and RT: Ymmetry Alance CalemashhoodBelum ada peringkat
- Choosing Sewer Safer Trees (SP628) PDFDokumen4 halamanChoosing Sewer Safer Trees (SP628) PDFMaria ComanBelum ada peringkat
- Problem SolvingDokumen2 halamanProblem SolvingmashhoodBelum ada peringkat
- 56192Dokumen25 halaman56192mashhoodBelum ada peringkat
- Power Management of A Stand-Alone Wind/Photovoltaic/Fuel Cell Energy SystemDokumen11 halamanPower Management of A Stand-Alone Wind/Photovoltaic/Fuel Cell Energy SystemAzza AhmedBelum ada peringkat
- Beekeeping Basics (Small Handout) WWDokumen26 halamanBeekeeping Basics (Small Handout) WWklaudyulBelum ada peringkat
- Quranic Search EngineDokumen4 halamanQuranic Search EnginemashhoodBelum ada peringkat
- Plants For Dry ClimatesDokumen4 halamanPlants For Dry ClimatesdvskiwiBelum ada peringkat
- Fungus GnatsDokumen2 halamanFungus GnatsSharad BhutoriaBelum ada peringkat
- Free Piston Stirling 7kwDokumen8 halamanFree Piston Stirling 7kwmikialeBelum ada peringkat
- So You Want To Be A Farmer: Book v2 7/7/99 1:09 PM Page 1Dokumen60 halamanSo You Want To Be A Farmer: Book v2 7/7/99 1:09 PM Page 1Emerson Pessôa FerreiraBelum ada peringkat
- Surah 1 Ayat 1-7 TranslationDokumen1 halamanSurah 1 Ayat 1-7 TranslationmashhoodBelum ada peringkat
- 12 Steps To StartupDokumen10 halaman12 Steps To StartupmashhoodBelum ada peringkat
- Media Literacy McsDokumen11 halamanMedia Literacy McsbabushajanBelum ada peringkat
- The Rational Analytical Approach To Decision-Making: An Adequate Strategy For Military Commanders?Dokumen23 halamanThe Rational Analytical Approach To Decision-Making: An Adequate Strategy For Military Commanders?mashhoodBelum ada peringkat
- Introduction To Cognitive Science: 1 CrumDokumen10 halamanIntroduction To Cognitive Science: 1 CrummashhoodBelum ada peringkat
- Surah 1 Ayat 1-7 TranslationDokumen1 halamanSurah 1 Ayat 1-7 TranslationmashhoodBelum ada peringkat
- The Distinction Between God's Essence and Energy: Gregory Palamas' Idea of Ultimate Reality and MeaningDokumen27 halamanThe Distinction Between God's Essence and Energy: Gregory Palamas' Idea of Ultimate Reality and MeaningmashhoodBelum ada peringkat
- Araucaria: Software For Argument Analysis, Diagramming and RepresentationDokumen19 halamanAraucaria: Software For Argument Analysis, Diagramming and RepresentationmashhoodBelum ada peringkat
- Theory, Culture & Society: Religion And/as MediaDokumen28 halamanTheory, Culture & Society: Religion And/as MediamashhoodBelum ada peringkat
- Political DiversityDokumen8 halamanPolitical DiversitymashhoodBelum ada peringkat
- Analytic InductionDokumen19 halamanAnalytic InductionpmjcsBelum ada peringkat
- God ConfusionDokumen217 halamanGod ConfusionmashhoodBelum ada peringkat
- Thewordbrain MicroeditionDokumen1 halamanThewordbrain MicroeditionFazle RabbiBelum ada peringkat
- Modelling Wind TurbineDokumen16 halamanModelling Wind Turbineranjit82Belum ada peringkat
- Aqidah Al-Tahawiyya by Imam Abu Ja'far Al-Tahawi Al-Hanafi (239-321 AH) Translated by Iqbal Ahmad AzamiDokumen9 halamanAqidah Al-Tahawiyya by Imam Abu Ja'far Al-Tahawi Al-Hanafi (239-321 AH) Translated by Iqbal Ahmad AzamimashhoodBelum ada peringkat
- Glossary Social Studies TermsDokumen35 halamanGlossary Social Studies TermsAhmed El-SaadanyBelum ada peringkat
- Everything You Always Wanted To Know About: Explained by John Baber, A.B.,Cd.EDokumen7 halamanEverything You Always Wanted To Know About: Explained by John Baber, A.B.,Cd.EmashhoodBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (119)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Nervous System IIDokumen33 halamanNervous System IIGeorgette TouzetBelum ada peringkat
- Abdomen: Radiology RevisionDokumen11 halamanAbdomen: Radiology RevisionhimaniBelum ada peringkat
- Water Balance in Human (Report)Dokumen4 halamanWater Balance in Human (Report)Syairah87Belum ada peringkat
- Brain Parts and Their Functions in LearningDokumen6 halamanBrain Parts and Their Functions in LearningChabbigillBelum ada peringkat
- MCQs - CRF & BalanceDokumen2 halamanMCQs - CRF & BalanceSara RezaBelum ada peringkat
- Diencephalon Thalamus and EpithalamusDokumen19 halamanDiencephalon Thalamus and EpithalamusIgme Mocke StraussBelum ada peringkat
- Imunologi Kulit DahDokumen17 halamanImunologi Kulit DahSayyid Muzahar AL-idruzzBelum ada peringkat
- Human Physiology Final ProjectDokumen37 halamanHuman Physiology Final ProjectAyesha MasoodBelum ada peringkat
- Signs and Symptoms of Pancreatic Cancer Fact Sheet Dec 2014Dokumen6 halamanSigns and Symptoms of Pancreatic Cancer Fact Sheet Dec 2014Nus EuBelum ada peringkat
- Ineffective Breast FeedingDokumen5 halamanIneffective Breast Feedingcamilleandaj070575% (12)
- Fisiologi Pengaturan Tekanan Darah SILBERNAGL 2Dokumen6 halamanFisiologi Pengaturan Tekanan Darah SILBERNAGL 221701101016 - Juliana Ayu NugrahaBelum ada peringkat
- 4th Term - G8 Daily Test (Biology) - MSDokumen17 halaman4th Term - G8 Daily Test (Biology) - MSMatthew EdbertBelum ada peringkat
- Revived Bcis2 TrialDokumen10 halamanRevived Bcis2 TrialSundaresan SankarBelum ada peringkat
- Approach To Abnormal Liver Enzymes Whitepaper TI 07106Dokumen6 halamanApproach To Abnormal Liver Enzymes Whitepaper TI 07106luceroBelum ada peringkat
- Chapter 06Dokumen47 halamanChapter 06gayathmipereraBelum ada peringkat
- Cardiovascular Anatomy and FunctionsDokumen6 halamanCardiovascular Anatomy and FunctionsKerwin LimBelum ada peringkat
- Extrahepatic Biliary ObstructionDokumen44 halamanExtrahepatic Biliary ObstructionOssama Abd Al-amierBelum ada peringkat
- Basal GangliaDokumen29 halamanBasal Gangliaapi-19916399Belum ada peringkat
- Syllabus DresserDokumen2 halamanSyllabus DresserVikash Aggarwal50% (2)
- Althea NCM 116aDokumen192 halamanAlthea NCM 116aspain michaelisBelum ada peringkat
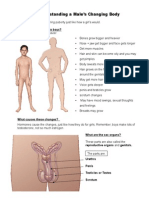
- 3 - Puberty Male Notes - TeacherDokumen3 halaman3 - Puberty Male Notes - Teacherapi-241585431100% (1)
- Master Radiology Notes GIT PDFDokumen115 halamanMaster Radiology Notes GIT PDFuroshkgBelum ada peringkat
- Medical Neuroscience Tutorial Notes: Overview of Neural SignalingDokumen2 halamanMedical Neuroscience Tutorial Notes: Overview of Neural SignalingOmar AlemánBelum ada peringkat
- Nervous System - All NotesDokumen6 halamanNervous System - All NotesJBelum ada peringkat
- What Is Rectum - 3Dokumen9 halamanWhat Is Rectum - 3Ela MarieBelum ada peringkat
- Benign Breast Conditions Malignant Breast Conditions: A: Breast Tissue. Connective Tissue, Fat TissueDokumen1 halamanBenign Breast Conditions Malignant Breast Conditions: A: Breast Tissue. Connective Tissue, Fat TissuedevilstBelum ada peringkat
- Pathophysiology: PneumoniaDokumen7 halamanPathophysiology: PneumoniaCindy Mae de la TorreBelum ada peringkat
- Normal Fetal Growth and DevelopmentDokumen38 halamanNormal Fetal Growth and DevelopmentArhum SiddiqiBelum ada peringkat
- Lecture: Adrenal DisordersDokumen78 halamanLecture: Adrenal DisordersOchendo KingxBelum ada peringkat
- TSH Conversion PDFDokumen1 halamanTSH Conversion PDFMarianBelum ada peringkat