Anda mungkin juga menyukai
- Embryology Article John WilksDokumen4 halamanEmbryology Article John Wilkspfi_jenBelum ada peringkat
- Trigger Point: A Luc Fortesque AdventureDari EverandTrigger Point: A Luc Fortesque AdventurePenilaian: 5 dari 5 bintang5/5 (1)
- Excerpts From "The Fasciae"Dokumen24 halamanExcerpts From "The Fasciae"intoilmariBelum ada peringkat
- Handbook of Us: Understanding and Accepting People with AutismDari EverandHandbook of Us: Understanding and Accepting People with AutismBelum ada peringkat
- The Bioenergetic Model in Osteopathic Diagnosis and Treatment: An FAAO Thesis, Part 1Dokumen9 halamanThe Bioenergetic Model in Osteopathic Diagnosis and Treatment: An FAAO Thesis, Part 1nish0420100% (1)
- A Model of Human Postural Control Inspired by Separated Human Sensory SystemsDokumen10 halamanA Model of Human Postural Control Inspired by Separated Human Sensory SystemspamelaBelum ada peringkat
- Bio Dynamics The Fascia of The Pelvic Floor Model of TensegrityDokumen8 halamanBio Dynamics The Fascia of The Pelvic Floor Model of TensegritygabrielsantosmsBelum ada peringkat
- The Fascia As Organ of Innerness 2015 EN ChapterDokumen16 halamanThe Fascia As Organ of Innerness 2015 EN ChapterJorge Flores TorresBelum ada peringkat
- Visceral - Manipulation A Powerful New Frontier in BodyworkDokumen5 halamanVisceral - Manipulation A Powerful New Frontier in BodyworkDerek Kim100% (1)
- Jim Jealous, - Healing and The Natural WorldDokumen8 halamanJim Jealous, - Healing and The Natural WorldSeda Joseph Saine100% (1)
- Plasticity FascialDokumen13 halamanPlasticity FascialMichael DuranBelum ada peringkat
- Aaron Driver Doctor of PhysiotherapyDokumen12 halamanAaron Driver Doctor of Physiotherapyaaronjdriver2248Belum ada peringkat
- Neuro AnatomyDokumen26 halamanNeuro Anatomymeryclave345Belum ada peringkat
- Sensorimotor Training A Global Approach For BalancDokumen9 halamanSensorimotor Training A Global Approach For BalancVizaBelum ada peringkat
- ReeducationSensorimotor PrinciplesandAModelinPhysiotherapyDokumen5 halamanReeducationSensorimotor PrinciplesandAModelinPhysiotherapyayuBelum ada peringkat
- Diagnostic Touch Its Principles and Application PDFDokumen9 halamanDiagnostic Touch Its Principles and Application PDFJakobStierleBelum ada peringkat
- Body Schema Building During Childhood and AdolescenceDokumen10 halamanBody Schema Building During Childhood and AdolescenceAída MoraBelum ada peringkat
- Chapmans Reflex 479Dokumen5 halamanChapmans Reflex 479Huram-abiBelum ada peringkat
- OPP Course Review: John Garlitz, DO Deborah Schmidt, DO Opp 2 April 15, 2015 8:10 AMDokumen248 halamanOPP Course Review: John Garlitz, DO Deborah Schmidt, DO Opp 2 April 15, 2015 8:10 AMmina botross100% (1)
- Lossing DiagnosingTreatingDu PDFDokumen33 halamanLossing DiagnosingTreatingDu PDFDiana SchlittlerBelum ada peringkat
- Physics of Cerebrospinal Fluid CirculationDokumen37 halamanPhysics of Cerebrospinal Fluid CirculationreaderwriteBelum ada peringkat
- Anatomi PelvisDokumen44 halamanAnatomi Pelvisari naBelum ada peringkat
- Chapman's Reflexes PointsDokumen10 halamanChapman's Reflexes PointscatalinBelum ada peringkat
- A Fascia and The Fascial SystemDokumen2 halamanA Fascia and The Fascial SystemGriBelum ada peringkat
- Physical Therapy Treatment of Pudendal Neuralgia: Elizabeth Rummer, MSPT Stephanie Prendergast, MPTDokumen30 halamanPhysical Therapy Treatment of Pudendal Neuralgia: Elizabeth Rummer, MSPT Stephanie Prendergast, MPTRamya Vasan100% (1)
- Fascia PDFDokumen68 halamanFascia PDFSteve67% (3)
- Sutherland's Legacy in The New MillenniumDokumen7 halamanSutherland's Legacy in The New MillenniumIsmael Vara CarvalhoBelum ada peringkat
- FasciaDokumen12 halamanFasciaRodolfo CiqueleiroBelum ada peringkat
- 2003 Fascial Plasticity - A New Neurobiological Explanation - Part 1 and Part 2Dokumen22 halaman2003 Fascial Plasticity - A New Neurobiological Explanation - Part 1 and Part 21Rogerio100% (1)
- SomaticsColorCatalog5 4 06Dokumen17 halamanSomaticsColorCatalog5 4 06Viviana PueblaBelum ada peringkat
- Head & Neck - Deep Cervical Fascia &cervical PlexusDokumen9 halamanHead & Neck - Deep Cervical Fascia &cervical PlexusKomang Tri Adi SuparwatiBelum ada peringkat
- Lossing BerylArbucklesCranial PDFDokumen67 halamanLossing BerylArbucklesCranial PDFdeemoney3100% (1)
- Understanding The Stretch ReflexDokumen2 halamanUnderstanding The Stretch ReflexJingyan ZhangBelum ada peringkat
- The Occipital BoneDokumen5 halamanThe Occipital Boneinna3003Belum ada peringkat
- NST Book enDokumen66 halamanNST Book enMiguelGutierrez100% (2)
- Sergueef N Nelson KE Osteopathy For TheDokumen12 halamanSergueef N Nelson KE Osteopathy For TheGabriela FerraciniBelum ada peringkat
- The Five Diaphragms in Osteopathic Manipulative MeDokumen14 halamanThe Five Diaphragms in Osteopathic Manipulative MeShaun Tyler100% (1)
- Clinical Examination of The Sacroiliac JointDokumen7 halamanClinical Examination of The Sacroiliac Jointthe_walnutBelum ada peringkat
- Cranial NervesDokumen16 halamanCranial Nervesapi-302547403100% (1)
- Visceral OMT: AAO Convocation March 2018 Kenneth Lossing DODokumen59 halamanVisceral OMT: AAO Convocation March 2018 Kenneth Lossing DODiana SchlittlerBelum ada peringkat
- Autonomic Nervous SystemDokumen41 halamanAutonomic Nervous SystemYanto HaryantoBelum ada peringkat
- The Cranial Bowl in The New Millennium and Sutherland's Legacy For Osteopathic Medicine: Part 2Dokumen8 halamanThe Cranial Bowl in The New Millennium and Sutherland's Legacy For Osteopathic Medicine: Part 2Juanillo CraneosacralBelum ada peringkat
- Osteopath yDokumen26 halamanOsteopath yManuel DamilBelum ada peringkat
- Gut and Brain Axis-Must READDokumen11 halamanGut and Brain Axis-Must READGenevieve JardineBelum ada peringkat
- Female Reproductive System Visceral ManipuationDokumen17 halamanFemale Reproductive System Visceral ManipuationFatemah Beyg100% (1)
- 2015 Inclusion of Trigger Point Dry Needling in A Multimodal Physical Therapy Program For Postoperative Shoulder Pain, A Randomized Clinical TrialDokumen9 halaman2015 Inclusion of Trigger Point Dry Needling in A Multimodal Physical Therapy Program For Postoperative Shoulder Pain, A Randomized Clinical TrialCarlos Meriño GuzmanBelum ada peringkat
- 1.temporal Bone & Skull Base Anatomy Seminar 1Dokumen67 halaman1.temporal Bone & Skull Base Anatomy Seminar 1fitsumalemayehudBelum ada peringkat
- The Clinical Applications of CranioSacral TherapyDokumen12 halamanThe Clinical Applications of CranioSacral Therapykabshiel100% (1)
- Eyal Lederman - Process Approach in PTDokumen72 halamanEyal Lederman - Process Approach in PTYuldash100% (1)
- Diagnostic Touch Parts I IV by Rollin Becker PDFDokumen69 halamanDiagnostic Touch Parts I IV by Rollin Becker PDFCatalin EneBelum ada peringkat
- Article Perfect Posture PuzzleDokumen6 halamanArticle Perfect Posture PuzzleMuhammad FahmyBelum ada peringkat
- The Cranial Bowl in The New Millennium and Sutherland's Legacy For Osteopathic Medicine: Part 1Dokumen9 halamanThe Cranial Bowl in The New Millennium and Sutherland's Legacy For Osteopathic Medicine: Part 1Juanillo CraneosacralBelum ada peringkat
- Neuro Estructural Integracion Tecnica PDF en InglesDokumen10 halamanNeuro Estructural Integracion Tecnica PDF en InglesLuulaa Minguin GarciaBelum ada peringkat
- Fascia Forgotten StructureDokumen12 halamanFascia Forgotten StructureFraan GutierrezBelum ada peringkat
- Essentials of o Ste 00 DaveDokumen112 halamanEssentials of o Ste 00 DaveGeki Iovan100% (1)
- Autonomic Nervous System: DR S S Ranasinghe CNTH - RagamaDokumen63 halamanAutonomic Nervous System: DR S S Ranasinghe CNTH - RagamawellawalalasithBelum ada peringkat
- Brain Based Gait Assessment FinalDokumen13 halamanBrain Based Gait Assessment FinalJESA MARI S SALONGABelum ada peringkat
- Fascial Strain Counter Strain 2013Dokumen8 halamanFascial Strain Counter Strain 2013Rui Pedro PereiraBelum ada peringkat
- Morin e Coll. Osteoptia Pediatrica Full PDFDokumen8 halamanMorin e Coll. Osteoptia Pediatrica Full PDFthe_walnutBelum ada peringkat
- Accuracy of Spinal Orthopaedic Tests: A Systematic ReviewDokumen10 halamanAccuracy of Spinal Orthopaedic Tests: A Systematic Reviewthe_walnutBelum ada peringkat
- Variations of High Frequency Parameter of Heart Rate Variability Following Osteopathic Manipulative TreatmentDokumen13 halamanVariations of High Frequency Parameter of Heart Rate Variability Following Osteopathic Manipulative Treatmentthe_walnutBelum ada peringkat
- An Alternative Approach To The Gastroesophageal Reflux Disease - Manual Techniques and NutritionDokumen10 halamanAn Alternative Approach To The Gastroesophageal Reflux Disease - Manual Techniques and Nutritionthe_walnutBelum ada peringkat
- Bio-Photons and Bio-Communication: Abstract-The Topic of Bio-Informational Aspects of Photon Emission Has ADokumen15 halamanBio-Photons and Bio-Communication: Abstract-The Topic of Bio-Informational Aspects of Photon Emission Has Aapi-271869539Belum ada peringkat
- Handbook of Osteopathic TechniqueDokumen60 halamanHandbook of Osteopathic Techniquethe_walnut100% (2)
- Clinical Examination of The Sacroiliac JointDokumen7 halamanClinical Examination of The Sacroiliac Jointthe_walnutBelum ada peringkat
- Common Biomechanical Dysfunctions Upper ExtremityDokumen10 halamanCommon Biomechanical Dysfunctions Upper Extremitythe_walnutBelum ada peringkat
- Handbook of Osteopathic TechniqueDokumen60 halamanHandbook of Osteopathic Techniquethe_walnut100% (2)
- Application of Derivative-04 - ExerciseDokumen13 halamanApplication of Derivative-04 - ExerciseRaju SinghBelum ada peringkat
- MA8151 Engineering Mathematics IDokumen14 halamanMA8151 Engineering Mathematics IRasheeda BeautifulBelum ada peringkat
- Advanced Data Structures and AlgorithmsDokumen7 halamanAdvanced Data Structures and AlgorithmsPrahitha MovvaBelum ada peringkat
- Calculus Value'able Theorems: Activity OverviewDokumen3 halamanCalculus Value'able Theorems: Activity OverviewGiova EleroseBelum ada peringkat
- Calculus of Management I Course Notes PDFDokumen270 halamanCalculus of Management I Course Notes PDFAriaBelum ada peringkat
- Lesson 25 Game TheoryDokumen25 halamanLesson 25 Game TheoryMousumi MajumdarBelum ada peringkat
- Bird - Higher Engineering Mathematics, 5TH Ed., Newnes, 2006Dokumen16 halamanBird - Higher Engineering Mathematics, 5TH Ed., Newnes, 2006RistagenBelum ada peringkat
- Mathematical OptimizationDokumen15 halamanMathematical OptimizationkentbnxBelum ada peringkat
- Closed Interval Method On eMathHelpDokumen1 halamanClosed Interval Method On eMathHelpShannon OrBelum ada peringkat
- CALCULUS PROTTER and PROTTERDokumen21 halamanCALCULUS PROTTER and PROTTERsilygierl100% (4)
- Lagrange's Equations PDFDokumen26 halamanLagrange's Equations PDFDebkanta DasBelum ada peringkat
- A Stability Function For Explicit Evaluation of The Mullins-Sekerka Interface Stability CriterionDokumen4 halamanA Stability Function For Explicit Evaluation of The Mullins-Sekerka Interface Stability CriterionKamel HebbacheBelum ada peringkat
- KAS 103 - Unit 3 - Lecture - 6Dokumen18 halamanKAS 103 - Unit 3 - Lecture - 6sagar guptaBelum ada peringkat
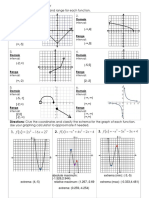
- Directions: Find The Domain and Range For Each FunctionDokumen3 halamanDirections: Find The Domain and Range For Each FunctionKayla IkumaBelum ada peringkat
- h111-h216 Hipot en EsDokumen118 halamanh111-h216 Hipot en Esjaved shaikh chaandBelum ada peringkat
- UNIT 2: Basic Concepts of Derivatives Lesson 7 - OptimizationDokumen20 halamanUNIT 2: Basic Concepts of Derivatives Lesson 7 - OptimizationJmBelum ada peringkat
- Lectures On Optimal Control Theory: Terje Sund March 3, 2014Dokumen54 halamanLectures On Optimal Control Theory: Terje Sund March 3, 2014عبدالرحمن الزهرانيBelum ada peringkat
- Algebra 2 - Polynomials - End BehaviorDokumen8 halamanAlgebra 2 - Polynomials - End BehaviorAYEL FARPARANBelum ada peringkat
- Mechleri, ChERD, 2016, RevisedDokumen30 halamanMechleri, ChERD, 2016, RevisedFitria Nur LailyBelum ada peringkat
- Ace Paper SolutionsDokumen10 halamanAce Paper SolutionsJack BaeBelum ada peringkat
- Lo, Mamaysky &wang - Foundations of Technical Analysis PDFDokumen61 halamanLo, Mamaysky &wang - Foundations of Technical Analysis PDFkhrysttalexa3146Belum ada peringkat
- (Jerome Spanier, Keith B. Oldham) An Atlas of Func PDFDokumen708 halaman(Jerome Spanier, Keith B. Oldham) An Atlas of Func PDFCarlos CasasolaBelum ada peringkat
- Mathematical Techniques: Revision Notes: DR A. J. BevanDokumen11 halamanMathematical Techniques: Revision Notes: DR A. J. BevanRoy VeseyBelum ada peringkat
- Managerial Economics - DerivativesDokumen4 halamanManagerial Economics - Derivativesjescy pauloBelum ada peringkat
- An Efficient 3D Toplogy Optimization Code Written in MATLABDokumen22 halamanAn Efficient 3D Toplogy Optimization Code Written in MATLABBarraq_00Belum ada peringkat
- Maths PDFDokumen43 halamanMaths PDFskg21813Belum ada peringkat
- Hsslive-12 LPP PDFDokumen4 halamanHsslive-12 LPP PDFTeam Electrical inspectorBelum ada peringkat
- Mathematics (Integral Caluculus) - Day 10: Maxima and MinimaDokumen14 halamanMathematics (Integral Caluculus) - Day 10: Maxima and MinimaRALPH JEAN CA�AREJOBelum ada peringkat
- Partial DerivativeDokumen71 halamanPartial Derivative9898171006Belum ada peringkat
- Math Practice Paper 5 QPDokumen9 halamanMath Practice Paper 5 QPANURAG YadavBelum ada peringkat