Anda mungkin juga menyukai
- Bottom Up: An Integrated Neurological and Cognitive Behavioural Book Which Addresses the Key Principles of Neuropsychotherapy. Five Important Emotional Parts of the Brain with Skills Training.Dari EverandBottom Up: An Integrated Neurological and Cognitive Behavioural Book Which Addresses the Key Principles of Neuropsychotherapy. Five Important Emotional Parts of the Brain with Skills Training.Belum ada peringkat
- Gunnar 2009 Stress and Early Brain DevelopmentDokumen6 halamanGunnar 2009 Stress and Early Brain DevelopmentRo Mi NaBelum ada peringkat
- Adolescent Brain Development: Implications for BehaviorDari EverandAdolescent Brain Development: Implications for BehaviorBelum ada peringkat
- Mentalization in Adult Attachment NarratDokumen20 halamanMentalization in Adult Attachment NarratHannah HolandaBelum ada peringkat
- Well-Being and Affective Style: Neural Substrates and Biobehavioural CorrelatesDokumen17 halamanWell-Being and Affective Style: Neural Substrates and Biobehavioural CorrelatesGustavo Adolfo DomínguezBelum ada peringkat
- CFJKDokumen30 halamanCFJKKiwiBelum ada peringkat
- Pechtel P11Dokumen16 halamanPechtel P11cfrl25Belum ada peringkat
- Villaneza Group 2 Set B Chapters PDFDokumen27 halamanVillaneza Group 2 Set B Chapters PDFZm YoungBelum ada peringkat
- What Is A Psychological Misconception? Moving Toward An Empirical AnswerDokumen11 halamanWhat Is A Psychological Misconception? Moving Toward An Empirical AnswerJuan SolanoBelum ada peringkat
- Neuropsychological Profile of Children Adolescents and Adults Experiencing Maltreatment A Meta AnalysisDokumen23 halamanNeuropsychological Profile of Children Adolescents and Adults Experiencing Maltreatment A Meta AnalysisjuanpaBelum ada peringkat
- An Investigation of The Influence of Emotional Factors On Learning in Physics InstructionDokumen20 halamanAn Investigation of The Influence of Emotional Factors On Learning in Physics InstructionNaillah SabaBelum ada peringkat
- Neural Correlates of Social Exclusion During Adolescence: Understanding The Distress of Peer RejectionDokumen15 halamanNeural Correlates of Social Exclusion During Adolescence: Understanding The Distress of Peer RejectionJoeBelum ada peringkat
- Psychometric Properties of A New ADHD Screening Questionnaire Parent Report On The Potential Underlying Explanation of Inattention in Their SchoolDokumen17 halamanPsychometric Properties of A New ADHD Screening Questionnaire Parent Report On The Potential Underlying Explanation of Inattention in Their SchoolIngrid DíazBelum ada peringkat
- Brain: Arrested Development: Early Prefrontal Lesions Impair The Maturation of Moral JudgementDokumen8 halamanBrain: Arrested Development: Early Prefrontal Lesions Impair The Maturation of Moral JudgementCarola ValdésBelum ada peringkat
- Methods of Neuropsychological Assessment EsDokumen32 halamanMethods of Neuropsychological Assessment EsHermod26AnimeBelum ada peringkat
- Belsky 2010Dokumen20 halamanBelsky 2010Anonymous XZ5YEmBelum ada peringkat
- PEDSchapter 2Dokumen50 halamanPEDSchapter 2Dian HaniBelum ada peringkat
- Neuroanatomia Funcional Machado 3 Ed 2014Dokumen34 halamanNeuroanatomia Funcional Machado 3 Ed 2014ThiagoSoaresBelum ada peringkat
- A-350 Key Studies in PsychologyDokumen9 halamanA-350 Key Studies in PsychologyGunjeet KaurBelum ada peringkat
- Flex Cog Com Meddra D Ans y DepreDokumen13 halamanFlex Cog Com Meddra D Ans y Deprethiagocasetta11Belum ada peringkat
- Fundamental Research: The Conduct of Agricultural ScienceDokumen9 halamanFundamental Research: The Conduct of Agricultural ScienceYemaneDibetaBelum ada peringkat
- Gamble, Moreau, Tippett, Addis (2018) - Specificity of Future Thinking in Depression - A Meta-Analysis - PreprintDokumen39 halamanGamble, Moreau, Tippett, Addis (2018) - Specificity of Future Thinking in Depression - A Meta-Analysis - PreprintMekhla PandeyBelum ada peringkat
- Cognition and Emotion Future Directions Dalgleish, TimDokumen13 halamanCognition and Emotion Future Directions Dalgleish, Timferreira.laurinda7995Belum ada peringkat
- 2011 Adhd Npsy-AssessmentDokumen15 halaman2011 Adhd Npsy-AssessmentRoberta Finamore de AraujoBelum ada peringkat
- Experimental Psychology Experiment Proposal Working Title: ExperimenterDokumen5 halamanExperimental Psychology Experiment Proposal Working Title: ExperimenterZymon Andrew MaquintoBelum ada peringkat
- Assessing Selective Sustained Attention in 3 - To 5-Year-Old Children - Evidence From A New ParadigmDokumen20 halamanAssessing Selective Sustained Attention in 3 - To 5-Year-Old Children - Evidence From A New ParadigmMACARENA VASQUEZ TORRESBelum ada peringkat
- Experience Effects On Brain Development Possible Contributions To PsychopathologyDokumen31 halamanExperience Effects On Brain Development Possible Contributions To PsychopathologyShirleuy GonçalvesBelum ada peringkat
- Six Major Psychological TheoriesDokumen22 halamanSix Major Psychological TheoriesJoh Na AlvaradoBelum ada peringkat
- Research Paper Social AnxietyDokumen5 halamanResearch Paper Social Anxietyafeasdvym100% (1)
- Neuropsychology Dissertation IdeasDokumen8 halamanNeuropsychology Dissertation IdeasHelpWithYourPaperAurora100% (1)
- Schonert-Reichl Et Al. MindUpDokumen15 halamanSchonert-Reichl Et Al. MindUpPeterBelum ada peringkat
- Neuropsychological Assessment of Cultrually and Linguistically DiDokumen24 halamanNeuropsychological Assessment of Cultrually and Linguistically DiphodoraBelum ada peringkat
- Biopsychosocial Model in Depression Revisited: Mauro Garcia-Toro, Iratxe AguirreDokumen9 halamanBiopsychosocial Model in Depression Revisited: Mauro Garcia-Toro, Iratxe AguirreayannaxemBelum ada peringkat
- Experimental Psychology Experiment Proposal Working Title: ExperimenterDokumen5 halamanExperimental Psychology Experiment Proposal Working Title: ExperimenterZymon Andrew MaquintoBelum ada peringkat
- Neuro AssignmentDokumen12 halamanNeuro AssignmentNancy DrewBelum ada peringkat
- Anderson 2014Dokumen16 halamanAnderson 2014Bárbara FernandesBelum ada peringkat
- The Development of Anxiety Disorders: Considering The Contributions of Attachment and Emotion RegulationDokumen15 halamanThe Development of Anxiety Disorders: Considering The Contributions of Attachment and Emotion RegulationJohannes CordeBelum ada peringkat
- Martin Psicofarmacología Infantil PDFDokumen814 halamanMartin Psicofarmacología Infantil PDFCarolina Salazar VásquezBelum ada peringkat
- Current Trends in PsychologyDokumen4 halamanCurrent Trends in PsychologyMuhammad AbidBelum ada peringkat
- Pediatric PsychopharmacologyDokumen814 halamanPediatric PsychopharmacologyLo Gri Palma100% (1)
- Articol SpecialitateDokumen31 halamanArticol SpecialitateMadaleenaBelum ada peringkat
- Neuroscience and Psychodynamic PsychologyDokumen4 halamanNeuroscience and Psychodynamic PsychologyImani ImaniBelum ada peringkat
- 10 1017@S0954579419000282 PDFDokumen11 halaman10 1017@S0954579419000282 PDFSamucyBelum ada peringkat
- The Development of Anxiety Disorders Considering TDokumen16 halamanThe Development of Anxiety Disorders Considering TAnanyaa RodricksBelum ada peringkat
- TMP D712Dokumen44 halamanTMP D712FrontiersBelum ada peringkat
- 2010 - Garner - Emotional Competence and Its InfluencesDokumen25 halaman2010 - Garner - Emotional Competence and Its InfluencesMargarita SantoyoBelum ada peringkat
- Positive Emotions in EducationDokumen15 halamanPositive Emotions in Educationmanuel alvarezBelum ada peringkat
- The Role of Neuropsychological AssessmentDokumen28 halamanThe Role of Neuropsychological AssessmentJovani Gonzales VillarealBelum ada peringkat
- Cornitive Control Go No Go TaskDokumen16 halamanCornitive Control Go No Go TaskclaudiaBelum ada peringkat
- Facing Puberty Associations Between Pubertal Development and Neural Responses To Affective Facil DisplaysDokumen9 halamanFacing Puberty Associations Between Pubertal Development and Neural Responses To Affective Facil Displayspsic.tatianebezerraBelum ada peringkat
- Perpectias Ddacticas Sobre La EmapndaaDokumen12 halamanPerpectias Ddacticas Sobre La EmapndaaEduardo OlarteBelum ada peringkat
- Effect of Video Games On Attention)Dokumen12 halamanEffect of Video Games On Attention)freddieBelum ada peringkat
- Research Paper EmotionsDokumen4 halamanResearch Paper EmotionsafeawckewBelum ada peringkat
- Notes For p5+6Dokumen13 halamanNotes For p5+6StefanBelum ada peringkat
- Child and Human DevelopmentDokumen4 halamanChild and Human Developmentmayette manuelBelum ada peringkat
- Consequentes e Antecedentes Da Regulação EmocionalDokumen15 halamanConsequentes e Antecedentes Da Regulação EmocionalRitaBelum ada peringkat
- Shared Neural Basis For The Exposure ToDokumen6 halamanShared Neural Basis For The Exposure Tosandrashalom2Belum ada peringkat
- Emotional Intelligence Research PaperDokumen4 halamanEmotional Intelligence Research Paperafedetbma100% (1)
- Research Paper On DepressionDokumen8 halamanResearch Paper On Depressionfveec9sx100% (1)
- Essay Mental HealthDokumen9 halamanEssay Mental HealthAaron LeeBelum ada peringkat
- Glosar Ro en NatoDokumen5 halamanGlosar Ro en Natomadsheep01Belum ada peringkat
- Creația Produselor VestimentareDokumen8 halamanCreația Produselor VestimentareCretu IleanaBelum ada peringkat
- Foreword To The Inner TreeDokumen22 halamanForeword To The Inner TreeCretu IleanaBelum ada peringkat
- Marine Selenee The Movement Family Const PDFDokumen205 halamanMarine Selenee The Movement Family Const PDFCretu Ileana100% (1)
- Curriculum Vitae Replace With First Name(s) Surname(s)Dokumen2 halamanCurriculum Vitae Replace With First Name(s) Surname(s)Valentina100% (1)
- Analiza Experimente ParapsihologiceDokumen27 halamanAnaliza Experimente ParapsihologiceCretu Ileana0% (1)
- Definition of Group CohesionDokumen3 halamanDefinition of Group CohesionCretu IleanaBelum ada peringkat
- Adaptive Strategies in Obtaining A New Transitional Identity SpaceDokumen15 halamanAdaptive Strategies in Obtaining A New Transitional Identity SpaceCretu IleanaBelum ada peringkat
- AnxietyDokumen1 halamanAnxietyCretu IleanaBelum ada peringkat
- Spiritual IntelligenceDokumen8 halamanSpiritual IntelligenceCretu IleanaBelum ada peringkat
- The Fractal TechniqueDokumen8 halamanThe Fractal TechniqueCretu IleanaBelum ada peringkat
- Terapii ExperiențialeDokumen14 halamanTerapii ExperiențialeCretu IleanaBelum ada peringkat
- 64-2 P. 15-18 PDFDokumen4 halaman64-2 P. 15-18 PDFCretu IleanaBelum ada peringkat
- Cunningham CH1 PPT ESSDokumen19 halamanCunningham CH1 PPT ESSCretu IleanaBelum ada peringkat
- Metanarrative Exercise PDFDokumen2 halamanMetanarrative Exercise PDFCretu IleanaBelum ada peringkat
- 7 Theories of Human Nature PDFDokumen9 halaman7 Theories of Human Nature PDFCretu IleanaBelum ada peringkat
- Threat-Related Attentional Bias in Anxious and Nonanxious Individuals: A Meta-Analytic StudyDokumen24 halamanThreat-Related Attentional Bias in Anxious and Nonanxious Individuals: A Meta-Analytic StudySara HeBelum ada peringkat
- Stout Shackman Larson E2015-LibreDokumen7 halamanStout Shackman Larson E2015-LibreCretu IleanaBelum ada peringkat
- Anxiety Selectively Disrupts Visuospatial Working Memory-LibreDokumen22 halamanAnxiety Selectively Disrupts Visuospatial Working Memory-LibreCretu IleanaBelum ada peringkat
- Jcompoziţia Bichimică A Fructelor de Juglans RegiaDokumen5 halamanJcompoziţia Bichimică A Fructelor de Juglans RegiaCretu IleanaBelum ada peringkat
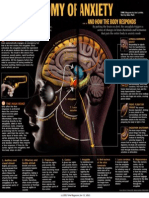
- Anatomy of AnxietyDokumen1 halamanAnatomy of AnxietyCretu IleanaBelum ada peringkat
- 2009 - Bio-Tex - Research Roadmap - March 2009Dokumen12 halaman2009 - Bio-Tex - Research Roadmap - March 2009Cretu IleanaBelum ada peringkat
- Clinical Assessment and Diagnosis: 1) Introduction 2) Basic Elements in Assessment 3) Physical AssessmentDokumen36 halamanClinical Assessment and Diagnosis: 1) Introduction 2) Basic Elements in Assessment 3) Physical AssessmentTrixieCamposano100% (1)
- Caffeine and Primary (Migraine) Headaches-Friend or Foe?: Karl B. Alstadhaug and Anna P. AndreouDokumen13 halamanCaffeine and Primary (Migraine) Headaches-Friend or Foe?: Karl B. Alstadhaug and Anna P. AndreouPutri SantriBelum ada peringkat
- Neuro Muscular Junction - KDokumen21 halamanNeuro Muscular Junction - Kkunjbihari sulakhiyaBelum ada peringkat
- Quiz #1 Q3 Science 10Dokumen2 halamanQuiz #1 Q3 Science 10Jr Capanang0% (1)
- The Happiness Advantage by Shawn AchorDokumen11 halamanThe Happiness Advantage by Shawn AchorsimasBelum ada peringkat
- Biomechanical Basis of Traumatic Brain InjuryDokumen32 halamanBiomechanical Basis of Traumatic Brain InjuryPutu AnantaBelum ada peringkat
- Maslow's Hierarchy of NeedsDokumen10 halamanMaslow's Hierarchy of NeedsMelissa FabillarBelum ada peringkat
- Paul Rhines - Eye Accessing Cues Mastery - Vol. 1 - Vol. 3 - WorkbookDokumen26 halamanPaul Rhines - Eye Accessing Cues Mastery - Vol. 1 - Vol. 3 - WorkbookC Damion HarperBelum ada peringkat
- Reception Learning David AusubelDokumen10 halamanReception Learning David AusubelKIM GED ALCOBERESBelum ada peringkat
- Assessment of Activities of Daily Living, Self-Care, and IndependenceDokumen11 halamanAssessment of Activities of Daily Living, Self-Care, and IndependenceayuBelum ada peringkat
- Approach To AtaxiaDokumen6 halamanApproach To AtaxiaVivek KumarBelum ada peringkat
- Membrane PotentialsDokumen40 halamanMembrane PotentialsdehaBelum ada peringkat
- Bansal V Et Al, 2006Dokumen7 halamanBansal V Et Al, 2006TriBelum ada peringkat
- Task-Based LearningDokumen9 halamanTask-Based LearningKatherine MondaBelum ada peringkat
- Vistas 2011 Article 53Dokumen13 halamanVistas 2011 Article 53Ateh HashimBelum ada peringkat
- Rancho Los Amigos RevDokumen3 halamanRancho Los Amigos RevSlvtierra Azu100% (1)
- Alcoholism Is A Disinhibitory Disorder: Neurophysiological Evidence From A Go/No-Go TaskDokumen21 halamanAlcoholism Is A Disinhibitory Disorder: Neurophysiological Evidence From A Go/No-Go TaskmirymirelaBelum ada peringkat
- Pemberton Happiness Index PDFDokumen13 halamanPemberton Happiness Index PDFIesanu MaraBelum ada peringkat
- TuriyaDokumen10 halamanTuriyaMircea CrivoiBelum ada peringkat
- Daftar Pustaka - 1Dokumen3 halamanDaftar Pustaka - 1Endang PasuluBelum ada peringkat
- Chapter 6Dokumen74 halamanChapter 6Umut Can TürkerBelum ada peringkat
- CHAPTER 5 OPERANT LEARNING FinalDokumen7 halamanCHAPTER 5 OPERANT LEARNING FinalWeyvin Maury Onde100% (1)
- 2007 - All That Glitters Is Not Gold' Misdiagnosis of Psychosis in Pervasive Developmental Disorders - A Case SeriesDokumen13 halaman2007 - All That Glitters Is Not Gold' Misdiagnosis of Psychosis in Pervasive Developmental Disorders - A Case Seriesamarillonoexpecta100% (1)
- Reading Microskills and Procedure For Training ThemDokumen11 halamanReading Microskills and Procedure For Training ThemtrinhquangcoBelum ada peringkat
- Grief and LossDokumen38 halamanGrief and LossMuarine Faye LaconsBelum ada peringkat
- Deficient Diversional ActivityDokumen2 halamanDeficient Diversional ActivityKimsha Concepcion100% (2)
- The Neuroscience of ReadingDokumen16 halamanThe Neuroscience of Readingapi-242291532Belum ada peringkat
- Dementia Types - Symptoms, Diagnosis, Causes, TreatmentsDokumen6 halamanDementia Types - Symptoms, Diagnosis, Causes, TreatmentsRatnaPrasadNalamBelum ada peringkat
- Preguntas Primer ExamenDokumen5 halamanPreguntas Primer ExamenMRVBelum ada peringkat
- Self-Hypnosis and Mental Imagery: PurposesDokumen7 halamanSelf-Hypnosis and Mental Imagery: PurposesTomas1122Belum ada peringkat
- The 7 Habits of Highly Effective People: The Infographics EditionDari EverandThe 7 Habits of Highly Effective People: The Infographics EditionPenilaian: 4 dari 5 bintang4/5 (2475)
- The Stoic Mindset: Living the Ten Principles of StoicismDari EverandThe Stoic Mindset: Living the Ten Principles of StoicismPenilaian: 4.5 dari 5 bintang4.5/5 (10)
- Summary: Atomic Habits by James Clear: An Easy & Proven Way to Build Good Habits & Break Bad OnesDari EverandSummary: Atomic Habits by James Clear: An Easy & Proven Way to Build Good Habits & Break Bad OnesPenilaian: 5 dari 5 bintang5/5 (1635)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeDari EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BePenilaian: 2 dari 5 bintang2/5 (1)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionDari EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionPenilaian: 4 dari 5 bintang4/5 (404)
- The Power of Now: A Guide to Spiritual EnlightenmentDari EverandThe Power of Now: A Guide to Spiritual EnlightenmentPenilaian: 4.5 dari 5 bintang4.5/5 (4125)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDari EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityPenilaian: 4 dari 5 bintang4/5 (3)
- Speaking Effective English!: Your Guide to Acquiring New Confidence In Personal and Professional CommunicationDari EverandSpeaking Effective English!: Your Guide to Acquiring New Confidence In Personal and Professional CommunicationPenilaian: 4.5 dari 5 bintang4.5/5 (74)
- Master Your Emotions: Develop Emotional Intelligence and Discover the Essential Rules of When and How to Control Your FeelingsDari EverandMaster Your Emotions: Develop Emotional Intelligence and Discover the Essential Rules of When and How to Control Your FeelingsPenilaian: 4.5 dari 5 bintang4.5/5 (321)
- Indistractable: How to Control Your Attention and Choose Your LifeDari EverandIndistractable: How to Control Your Attention and Choose Your LifePenilaian: 3 dari 5 bintang3/5 (5)
- The One Thing: The Surprisingly Simple Truth Behind Extraordinary ResultsDari EverandThe One Thing: The Surprisingly Simple Truth Behind Extraordinary ResultsPenilaian: 4.5 dari 5 bintang4.5/5 (709)
- Becoming Supernatural: How Common People Are Doing The UncommonDari EverandBecoming Supernatural: How Common People Are Doing The UncommonPenilaian: 5 dari 5 bintang5/5 (1480)
- Summary: The Laws of Human Nature: by Robert Greene: Key Takeaways, Summary & AnalysisDari EverandSummary: The Laws of Human Nature: by Robert Greene: Key Takeaways, Summary & AnalysisPenilaian: 4.5 dari 5 bintang4.5/5 (30)
- The 5 Second Rule: Transform your Life, Work, and Confidence with Everyday CourageDari EverandThe 5 Second Rule: Transform your Life, Work, and Confidence with Everyday CouragePenilaian: 5 dari 5 bintang5/5 (9)
- The Science of Self Discipline: How Daily Self-Discipline, Everyday Habits and an Optimised Belief System will Help You Beat Procrastination + Why Discipline Equals True FreedomDari EverandThe Science of Self Discipline: How Daily Self-Discipline, Everyday Habits and an Optimised Belief System will Help You Beat Procrastination + Why Discipline Equals True FreedomPenilaian: 4.5 dari 5 bintang4.5/5 (867)
- The 16 Undeniable Laws of Communication: Apply Them and Make the Most of Your MessageDari EverandThe 16 Undeniable Laws of Communication: Apply Them and Make the Most of Your MessagePenilaian: 5 dari 5 bintang5/5 (73)
- Take Charge of Your Life: The Winner's SeminarDari EverandTake Charge of Your Life: The Winner's SeminarPenilaian: 5 dari 5 bintang5/5 (177)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDari EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisPenilaian: 3.5 dari 5 bintang3.5/5 (2)
- The Silva Mind Method: for Getting Help from the Other SideDari EverandThe Silva Mind Method: for Getting Help from the Other SidePenilaian: 5 dari 5 bintang5/5 (51)
- Gut: the new and revised Sunday Times bestsellerDari EverandGut: the new and revised Sunday Times bestsellerPenilaian: 4 dari 5 bintang4/5 (393)
- Mastering Productivity: Everything You Need to Know About Habit FormationDari EverandMastering Productivity: Everything You Need to Know About Habit FormationPenilaian: 4.5 dari 5 bintang4.5/5 (23)
- Own Your Past Change Your Future: A Not-So-Complicated Approach to Relationships, Mental Health & WellnessDari EverandOwn Your Past Change Your Future: A Not-So-Complicated Approach to Relationships, Mental Health & WellnessPenilaian: 5 dari 5 bintang5/5 (85)
- Quantum Success: 7 Essential Laws for a Thriving, Joyful, and Prosperous Relationship with Work and MoneyDari EverandQuantum Success: 7 Essential Laws for a Thriving, Joyful, and Prosperous Relationship with Work and MoneyPenilaian: 5 dari 5 bintang5/5 (38)
- The Motive: Why So Many Leaders Abdicate Their Most Important ResponsibilitiesDari EverandThe Motive: Why So Many Leaders Abdicate Their Most Important ResponsibilitiesPenilaian: 5 dari 5 bintang5/5 (224)
- Uptime: A Practical Guide to Personal Productivity and WellbeingDari EverandUptime: A Practical Guide to Personal Productivity and WellbeingBelum ada peringkat
- The Miracle Morning by Hal Elrod: A Summary and AnalysisDari EverandThe Miracle Morning by Hal Elrod: A Summary and AnalysisPenilaian: 4.5 dari 5 bintang4.5/5 (55)