Anda mungkin juga menyukai
- Cover Modul FCPDokumen3 halamanCover Modul FCPYusuf Almalik SaputraBelum ada peringkat
- 4 Vol. 7 Issue 1 Jan 2016 IJPSR RE 1738 Paper 4Dokumen11 halaman4 Vol. 7 Issue 1 Jan 2016 IJPSR RE 1738 Paper 4Yusuf Almalik SaputraBelum ada peringkat
- Uji Aktifitas Antikanker Ekstrak Etanol Kulit Batang Bruguiera Gymnorhiza Terhadap Sel Meyloma Warsinah, Puji Lestari, TrisnowatiDokumen7 halamanUji Aktifitas Antikanker Ekstrak Etanol Kulit Batang Bruguiera Gymnorhiza Terhadap Sel Meyloma Warsinah, Puji Lestari, TrisnowatiYusuf Almalik SaputraBelum ada peringkat
- Biochemistry of The EyeDokumen38 halamanBiochemistry of The EyeYusuf Almalik SaputraBelum ada peringkat
- Baby Blues & Postpartum Depression PDFDokumen2 halamanBaby Blues & Postpartum Depression PDFYusuf Almalik SaputraBelum ada peringkat
- Efektifitas Ekstrak Biji Srikaya (Annona Squamosa L) Dengan Pelarut Air, Metanol Dan Heksan Terhadap Mortalitas Larva Caplak Boophilus Microplus Secara in VitroDokumen9 halamanEfektifitas Ekstrak Biji Srikaya (Annona Squamosa L) Dengan Pelarut Air, Metanol Dan Heksan Terhadap Mortalitas Larva Caplak Boophilus Microplus Secara in VitroYusuf Almalik SaputraBelum ada peringkat
- Hormon Reproduksi: Dr. Septi Handayani, M.SiDokumen133 halamanHormon Reproduksi: Dr. Septi Handayani, M.SiYusuf Almalik SaputraBelum ada peringkat
- Kul. 6 Respiratory EmegencyDokumen46 halamanKul. 6 Respiratory EmegencyYusuf Almalik SaputraBelum ada peringkat
- Drugs Used in Gastrointestinal DiseasesDokumen44 halamanDrugs Used in Gastrointestinal DiseasesYusuf Almalik Saputra0% (1)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Neural Tube DefectsDokumen2 halamanNeural Tube DefectsVijay SluBelum ada peringkat
- Storage ConditionDokumen4 halamanStorage Conditioneshwar_1953Belum ada peringkat
- KVG Medical CollegeDokumen11 halamanKVG Medical CollegepentagoneducationBelum ada peringkat
- Brodil LevoDokumen2 halamanBrodil LevoAziza KhairunBelum ada peringkat
- 2010 Endocrine System ChartDokumen3 halaman2010 Endocrine System ChartAlexander QianBelum ada peringkat
- High Lateral Tension AbdominoplastyDokumen14 halamanHigh Lateral Tension AbdominoplastyAhmad Arbi AninditoBelum ada peringkat
- Anatomy and Physiology Respiratory SystemDokumen7 halamanAnatomy and Physiology Respiratory Systemnoronisa talusobBelum ada peringkat
- Thorax and Lungs SGDokumen2 halamanThorax and Lungs SGDestinee Caple100% (1)
- Navjaat Shishu Suraksha Karyakram: Training ManualDokumen99 halamanNavjaat Shishu Suraksha Karyakram: Training ManualBiswaranjan Padhy100% (1)
- Graseby 3100 Syringe Pump - Service ManualDokumen89 halamanGraseby 3100 Syringe Pump - Service ManualRamanjaneya goud100% (1)
- Oral Wound Healing Efficacy of 1% Myristica Fragrans (Nutmeg) Determined Using MTT Assay: An in Vitro StudyDokumen5 halamanOral Wound Healing Efficacy of 1% Myristica Fragrans (Nutmeg) Determined Using MTT Assay: An in Vitro StudyDeepthi ManjunathBelum ada peringkat
- TRAMADOLDokumen4 halamanTRAMADOLRudie Lee PascualBelum ada peringkat
- Limfoma Kutis Pada Pasien Yang Semula Di DiagnosisDokumen34 halamanLimfoma Kutis Pada Pasien Yang Semula Di DiagnosisChris MulyoBelum ada peringkat
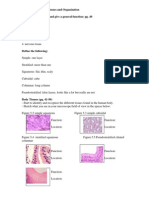
- Human Biology Laboratory 4: Body Tissues and Organization List The 4 Tissue Types and Give A General Function: Pg. 40Dokumen4 halamanHuman Biology Laboratory 4: Body Tissues and Organization List The 4 Tissue Types and Give A General Function: Pg. 40bugyourselfBelum ada peringkat
- Vasoactive Agents in Shock.2Dokumen8 halamanVasoactive Agents in Shock.2Linamaria LozanoBelum ada peringkat
- UKA: When Would I Do It?Dokumen35 halamanUKA: When Would I Do It?neareastspineBelum ada peringkat
- Soccer Programme U19 Pre in SeasonDokumen38 halamanSoccer Programme U19 Pre in SeasonJeffrey Peeko100% (1)
- Smear Layer Final2Dokumen10 halamanSmear Layer Final2bhudentBelum ada peringkat
- High Power Electrosurgery Review Update 2005Dokumen33 halamanHigh Power Electrosurgery Review Update 2005Ana ObradovicBelum ada peringkat
- 2018 - Vitamine D Treatment Guidelines in India - JClinSciResDokumen10 halaman2018 - Vitamine D Treatment Guidelines in India - JClinSciResnagashayana gBelum ada peringkat
- SialodenosisDokumen76 halamanSialodenosisJessica GreenBelum ada peringkat
- Drug Therapy in Pregnancy (Soal)Dokumen6 halamanDrug Therapy in Pregnancy (Soal)Yola FebriyantiBelum ada peringkat
- Cesarean Surgical Techniques - GLOWMDokumen36 halamanCesarean Surgical Techniques - GLOWMOana BalintBelum ada peringkat
- Role, Prospects, and Limitation of HomoeopathyDokumen43 halamanRole, Prospects, and Limitation of HomoeopathyDr.bhagyaBelum ada peringkat
- Neck Pain and Lower Back Pain. Med Clin N Am. 2019.Dokumen14 halamanNeck Pain and Lower Back Pain. Med Clin N Am. 2019.Jose Fernando Diez100% (1)
- Massive Transfusion ProtocolDokumen2 halamanMassive Transfusion ProtocolmukriBelum ada peringkat
- SCOLIOSISDokumen4 halamanSCOLIOSISapi-3822433Belum ada peringkat
- Efficacy and Adverse Events of Oral Isotretinoin For Acne: A Systematic ReviewDokumen10 halamanEfficacy and Adverse Events of Oral Isotretinoin For Acne: A Systematic ReviewFerryGoBelum ada peringkat
- At-Risk and Runaway Youth in Washington State: Outcomes For Youth Admitted To Secure Crisis Residential Centers and Mandatory Chemical Dependency TreatmentDokumen45 halamanAt-Risk and Runaway Youth in Washington State: Outcomes For Youth Admitted To Secure Crisis Residential Centers and Mandatory Chemical Dependency TreatmentWashington State Institute for Public PolicyBelum ada peringkat
- Cardiac GlycosidesDokumen8 halamanCardiac GlycosidesShan Sicat100% (1)