Anda mungkin juga menyukai
- The APBC Book of Companion Animal BehaviourDari EverandThe APBC Book of Companion Animal BehaviourPenilaian: 2 dari 5 bintang2/5 (2)
- Social Organization of The CatDokumen10 halamanSocial Organization of The CatElena PăunBelum ada peringkat
- Understanding Cats.: Compendium (Yardley, PA) May 2007Dokumen4 halamanUnderstanding Cats.: Compendium (Yardley, PA) May 2007CainBelum ada peringkat
- Primate Sociality and Social SystemsDokumen6 halamanPrimate Sociality and Social Systemsali haider bukhariBelum ada peringkat
- Pup Rearing: The Role of Mothers and Allomothers in Free-Ranging Domestic DogsDokumen13 halamanPup Rearing: The Role of Mothers and Allomothers in Free-Ranging Domestic DogsLucy LinBelum ada peringkat
- Aggressive Behaviors in Domestic CatsDokumen8 halamanAggressive Behaviors in Domestic CatsElida LiraBelum ada peringkat
- Group 9 Case 2.3Dokumen9 halamanGroup 9 Case 2.3aisyaa dwiBelum ada peringkat
- Dog Social Behavior and Communication: John Bradshaw and Nicola RooneyDokumen27 halamanDog Social Behavior and Communication: John Bradshaw and Nicola Rooneyalephe.leoneuBelum ada peringkat
- Aa FiveDokumen2 halamanAa FiveJennylyn BlaquirBelum ada peringkat
- (Felis Silvestris Cams) : The Function of Allogrooming in Domestic CatsDokumen13 halaman(Felis Silvestris Cams) : The Function of Allogrooming in Domestic CatsMarkBelum ada peringkat
- A Review of Environmental Enrichment For Kennelled Dogs, Canis FamiliarisDokumen11 halamanA Review of Environmental Enrichment For Kennelled Dogs, Canis Familiarisrazan almoajelBelum ada peringkat
- Dogs As Human Companions: A Review of The Relationship: January 1995Dokumen28 halamanDogs As Human Companions: A Review of The Relationship: January 1995Vladut BratfaleanBelum ada peringkat
- Environmental Aspects of Domestic Cat Care and ManagementDokumen8 halamanEnvironmental Aspects of Domestic Cat Care and ManagementAllyssa ReadsBelum ada peringkat
- Literature Rev Final DraftDokumen14 halamanLiterature Rev Final Draftapi-385926661Belum ada peringkat
- Simios SimiosDokumen18 halamanSimios SimiosNikaBelum ada peringkat
- On The Effects of Domestication On Canine Social Development and BehaviorDokumen19 halamanOn The Effects of Domestication On Canine Social Development and Behaviorapi-583138028Belum ada peringkat
- Wolf Status and Dominance in Packs - Alpha StatusDokumen13 halamanWolf Status and Dominance in Packs - Alpha StatusShoshannah ForbesBelum ada peringkat
- Social Networks Reveal Sex - and Age-Patterned Social Structure in Butler's Gartersnakes (Thamnophis Butleri)Dokumen14 halamanSocial Networks Reveal Sex - and Age-Patterned Social Structure in Butler's Gartersnakes (Thamnophis Butleri)Frederick NakosBelum ada peringkat
- 35 Crowding Into The Behavioral Sink2Dokumen5 halaman35 Crowding Into The Behavioral Sink2dennis ridenourBelum ada peringkat
- Social BehaviorDokumen32 halamanSocial BehaviorNoor ul aminBelum ada peringkat
- HE Elfless ENE: Olivia JudsonDokumen6 halamanHE Elfless ENE: Olivia JudsonDuong NguyenBelum ada peringkat
- Department of Psychology, Boioling Green State University, Bowling Green, OhioDokumen10 halamanDepartment of Psychology, Boioling Green State University, Bowling Green, OhioHazen Dominique Caparros MaquintoBelum ada peringkat
- What's Inside Your Cat's Head? A Review of Cat (Felis Silvestris Catus) Cognition Research Past, Present and FutureDokumen32 halamanWhat's Inside Your Cat's Head? A Review of Cat (Felis Silvestris Catus) Cognition Research Past, Present and FutureRomina DatuBelum ada peringkat
- Degree of Behavioral Neoteny Differentiates Canids Polymorphs - ArchivezDokumen24 halamanDegree of Behavioral Neoteny Differentiates Canids Polymorphs - ArchivezLydia Amate AmaroBelum ada peringkat
- Ali Hassan (Ent-514)Dokumen18 halamanAli Hassan (Ent-514)Ali hassanBelum ada peringkat
- Minimalist Slides For MeetingyDokumen11 halamanMinimalist Slides For MeetingyahmedBelum ada peringkat
- Fabre-Nys - 2007 - Sexual Behaviour in Ewes and Other RuminantsDokumen8 halamanFabre-Nys - 2007 - Sexual Behaviour in Ewes and Other RuminantsGustavo Campos Bueno DiasBelum ada peringkat
- Pattern of Speciation of Felidae Zia Czarina A. GarciaDokumen14 halamanPattern of Speciation of Felidae Zia Czarina A. GarciaSedsed QuematonBelum ada peringkat
- Cooperative Breeding: Michael A. CantDokumen6 halamanCooperative Breeding: Michael A. CantDanna Estefania Lopez HernandezBelum ada peringkat
- Animal Behavior NoteDokumen27 halamanAnimal Behavior NoteAyalew DemekBelum ada peringkat
- HCP Draft-1Dokumen8 halamanHCP Draft-1api-322547661Belum ada peringkat
- Eusocial BehaviourDokumen24 halamanEusocial Behaviouralka nokhwalBelum ada peringkat
- Final Presentation On Animal CommunicationDokumen15 halamanFinal Presentation On Animal CommunicationDwayne Coelho100% (2)
- Johnson 2000Dokumen26 halamanJohnson 2000IASA73Belum ada peringkat
- More Than Human Families Pets People and PDFDokumen13 halamanMore Than Human Families Pets People and PDFsegataufrnBelum ada peringkat
- Behavioural Reactions of Elephants Towards A Dying MatriarchDokumen16 halamanBehavioural Reactions of Elephants Towards A Dying MatriarchFlorencia AstorgaBelum ada peringkat
- ScienceDokumen2 halamanScienceAlexa GraganzaBelum ada peringkat
- Minimalist Slides For MeetingDokumen11 halamanMinimalist Slides For MeetingahmedBelum ada peringkat
- Sociability: Three Cats Engage inDokumen2 halamanSociability: Three Cats Engage inqwerty aBelum ada peringkat
- Krutzen Et AlDokumen6 halamanKrutzen Et Alapi-3828346Belum ada peringkat
- Pummelish LionDokumen2 halamanPummelish LionThang ThangBelum ada peringkat
- Unit 1 EvaluationDokumen15 halamanUnit 1 Evaluationahmed.adel.mazen2000Belum ada peringkat
- Read CrowdingDokumen5 halamanRead Crowdingdennis ridenourBelum ada peringkat
- Behaviour Problems and The Domestic Rabbit - Apbc ChapterDokumen11 halamanBehaviour Problems and The Domestic Rabbit - Apbc ChaptertherealosaisBelum ada peringkat
- ArmitageDokumen11 halamanArmitagec4422716Belum ada peringkat
- Teamworking Unit 1Dokumen11 halamanTeamworking Unit 1Đô Đặng Lê ThànhBelum ada peringkat
- Aggressive Behavior of Dogs Kept As Companion Animals: Classification and Influence of Sex, Reproductive Status and BreedDokumen17 halamanAggressive Behavior of Dogs Kept As Companion Animals: Classification and Influence of Sex, Reproductive Status and BreedcBelum ada peringkat
- Pathways rw3 2e U1 TestDokumen13 halamanPathways rw3 2e U1 Testdieuanh100% (1)
- Pathways Reading, Writing, and Critical Thinking 2e: Level 3 Unit 1 Test Name: - Class: - DateDokumen7 halamanPathways Reading, Writing, and Critical Thinking 2e: Level 3 Unit 1 Test Name: - Class: - DateBinh Pham Thanh100% (1)
- Shah Et Al 2003 (Sociality in Lizards) BehaviourDokumen14 halamanShah Et Al 2003 (Sociality in Lizards) BehaviourjavieromaBelum ada peringkat
- Case 2008Dokumen7 halamanCase 2008José Manuel PeñaBelum ada peringkat
- Raising Animals To Die For Profit - How Corporate America Is ChangDokumen10 halamanRaising Animals To Die For Profit - How Corporate America Is ChangElmira HokmabadiBelum ada peringkat
- Altruism in Animal CommunitiesDokumen40 halamanAltruism in Animal CommunitiesWalghBelum ada peringkat
- OooooogggosDokumen2 halamanOooooogggosapi-236686558Belum ada peringkat
- Jus ADokumen7 halamanJus AOctavio GilBelum ada peringkat
- NPWRC Alpha Status Dominance and Division of Labor 1Dokumen2 halamanNPWRC Alpha Status Dominance and Division of Labor 1api-238976508Belum ada peringkat
- Functional Properties of Ant Queen Pheromones As Revealed by Behavioral ExperimentsDokumen21 halamanFunctional Properties of Ant Queen Pheromones As Revealed by Behavioral ExperimentsJosip PrimoracBelum ada peringkat
- Social Organisation in Animals.: Saju J Zacharia 17-PZO-017Dokumen29 halamanSocial Organisation in Animals.: Saju J Zacharia 17-PZO-017Saju John ZachariaBelum ada peringkat
- Mammal Review - 2021 - Muller - A Review of The Social Behaviour of The Giraffe Giraffa Camelopardalis A Misunderstood ButDokumen15 halamanMammal Review - 2021 - Muller - A Review of The Social Behaviour of The Giraffe Giraffa Camelopardalis A Misunderstood Butaguilarodriguez48812Belum ada peringkat
- Adopting A Very Shy or Feral Cat June2019Dokumen3 halamanAdopting A Very Shy or Feral Cat June2019Luis Antonio Buitron RamirezBelum ada peringkat
- Separation, Confinement, or Noises:: What Is Scaring That Dog?Dokumen20 halamanSeparation, Confinement, or Noises:: What Is Scaring That Dog?Luis Antonio Buitron RamirezBelum ada peringkat
- AAFP and ISFM Feline Guidelines PDFDokumen12 halamanAAFP and ISFM Feline Guidelines PDFLuis Antonio Buitron RamirezBelum ada peringkat
- EG01 Caring For Your CatDokumen20 halamanEG01 Caring For Your CatRick SlickBelum ada peringkat
- Dog Training Methods Review PDFDokumen47 halamanDog Training Methods Review PDFLuis Antonio Buitron RamirezBelum ada peringkat
- Importance of Puppy Training For Future Behavior of The Dog-0008 PDFDokumen9 halamanImportance of Puppy Training For Future Behavior of The Dog-0008 PDFLuis Antonio Buitron RamirezBelum ada peringkat
- Importance of Puppy Training For Future Behavior of The Dog-0008Dokumen7 halamanImportance of Puppy Training For Future Behavior of The Dog-0008Luis Antonio Buitron RamirezBelum ada peringkat
- Aspects of Juvenile Fear PDFDokumen8 halamanAspects of Juvenile Fear PDFLuis Antonio Buitron RamirezBelum ada peringkat
- Please Tell Us About Your Dog's BehaviorDokumen1 halamanPlease Tell Us About Your Dog's BehaviorLuis Antonio Buitron RamirezBelum ada peringkat
- Please Tell Us About Your Cat's BehaviorDokumen1 halamanPlease Tell Us About Your Cat's BehaviorLuis Antonio Buitron RamirezBelum ada peringkat
- Monitoreo Ansiedad Por SeparaciónDokumen1 halamanMonitoreo Ansiedad Por SeparaciónLuis Antonio Buitron RamirezBelum ada peringkat
- Mcguire2018 PDFDokumen44 halamanMcguire2018 PDFLuis Antonio Buitron RamirezBelum ada peringkat
- Hart Et Al-2018-Veterinary Medicine and ScienceDokumen9 halamanHart Et Al-2018-Veterinary Medicine and Scienceamydaniels99Belum ada peringkat
- The Behavioral AssessmentDokumen5 halamanThe Behavioral AssessmentLuis Antonio Buitron RamirezBelum ada peringkat
- Clinical Formulation For Mental Health Nursing PracticeDokumen8 halamanClinical Formulation For Mental Health Nursing PracticeLuis Antonio Buitron RamirezBelum ada peringkat
- Separation Anxiety: Pamphlets For Pet ParentsDokumen3 halamanSeparation Anxiety: Pamphlets For Pet ParentsLuis Antonio Buitron RamirezBelum ada peringkat
- 098 Canine and Feline D+mentiaDokumen166 halaman098 Canine and Feline D+mentiaLuis Antonio Buitron RamirezBelum ada peringkat
- Mouth-Licking by Dogs As A Response To Emotional StimuliDokumen4 halamanMouth-Licking by Dogs As A Response To Emotional StimuliLuis Antonio Buitron RamirezBelum ada peringkat
- Factor Neurotrofico Derivado Del CerebroDokumen9 halamanFactor Neurotrofico Derivado Del CerebroLuis Antonio Buitron RamirezBelum ada peringkat
- Protocolo de Relajación - Segunda ParteDokumen2 halamanProtocolo de Relajación - Segunda ParteLuis Antonio Buitron RamirezBelum ada peringkat
- Fisica Del Modelo HidraulicoDokumen10 halamanFisica Del Modelo HidraulicoLuis Antonio Buitron RamirezBelum ada peringkat
- The Relationship Between Coat Color andDokumen16 halamanThe Relationship Between Coat Color andLuis Antonio Buitron RamirezBelum ada peringkat
- Examining Dog-Human Play PDFDokumen10 halamanExamining Dog-Human Play PDFLuis Antonio Buitron RamirezBelum ada peringkat
- Comparative Psychology, Ethology and Anmial BehaviorDokumen22 halamanComparative Psychology, Ethology and Anmial BehaviorLuis Antonio Buitron RamirezBelum ada peringkat
- Diverio Et Al. 2008Dokumen13 halamanDiverio Et Al. 2008Luis Antonio Buitron RamirezBelum ada peringkat
- Ethological Models and The Concept of DiriveDokumen6 halamanEthological Models and The Concept of DiriveLuis Antonio Buitron RamirezBelum ada peringkat
- Recorrido Historico de La Etologia 2007Dokumen12 halamanRecorrido Historico de La Etologia 2007CaBeBelum ada peringkat
- Completing The ExaminatioDokumen4 halamanCompleting The ExaminatioLuis Antonio Buitron RamirezBelum ada peringkat
- Dealing With Ambivalence - Farmers and Consumers Perceptions of Animal Welfare in LivestockDokumen18 halamanDealing With Ambivalence - Farmers and Consumers Perceptions of Animal Welfare in LivestockLuis Antonio Buitron RamirezBelum ada peringkat
- Bird Watching Checklist - Source: Minnesota Department of Natural ResourcesDokumen2 halamanBird Watching Checklist - Source: Minnesota Department of Natural ResourcesinforumdocsBelum ada peringkat
- HormoneDokumen68 halamanHormoneVelrones TampubolonBelum ada peringkat
- Spotlight On Endangered Animals: Name DateDokumen2 halamanSpotlight On Endangered Animals: Name Datebobnh100% (2)
- Lec.3 Development of The FaceDokumen25 halamanLec.3 Development of The FaceDentistryBelum ada peringkat
- Q2 Summ Test 1Dokumen24 halamanQ2 Summ Test 1Eden PingliBelum ada peringkat
- Gr5 Ibt Eng PracticeDokumen7 halamanGr5 Ibt Eng PracticeArmaan50% (2)
- Eugene MunroeDokumen3 halamanEugene MunroeAndrew ZaikinBelum ada peringkat
- Acinii SalivariDokumen5 halamanAcinii SalivariIo DobriBelum ada peringkat
- Muscular TissueDokumen49 halamanMuscular TissueSteph VeeBelum ada peringkat
- Flatworms 1Dokumen63 halamanFlatworms 1Budi AfriyansyahBelum ada peringkat
- Eukaryotic and Prokaryotic Gene StructureDokumen5 halamanEukaryotic and Prokaryotic Gene Structuredaniel murilloBelum ada peringkat
- Generus ClasificadorDokumen262 halamanGenerus Clasificadorchristopher suarezBelum ada peringkat
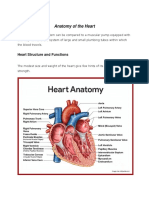
- Anatomy of The HeartDokumen18 halamanAnatomy of The Heartvanessaresullar100% (1)
- Animal Essay RevisedDokumen5 halamanAnimal Essay Revisedapi-315604985Belum ada peringkat
- Chordata NotesDokumen5 halamanChordata NotesAbhishek Isaac MathewBelum ada peringkat
- What To Protect? Systematics and The Agony of Choice: R. I. Vane-Wright, C. J. Humphries & P. H. WilliamsDokumen20 halamanWhat To Protect? Systematics and The Agony of Choice: R. I. Vane-Wright, C. J. Humphries & P. H. WilliamsLopa LopaBelum ada peringkat
- Animal Thematic 1852 - 2011: Tony BrayDokumen227 halamanAnimal Thematic 1852 - 2011: Tony Braydeepheat_008Belum ada peringkat
- MCAT 2014 Biology Portion Answer Key: WhiteDokumen14 halamanMCAT 2014 Biology Portion Answer Key: WhiteMuhammad ShahzadBelum ada peringkat
- Gill SlitDokumen2 halamanGill SlitNirmal BhowmickBelum ada peringkat
- Oxford Phonic Writing 01 PDFDokumen28 halamanOxford Phonic Writing 01 PDFNhợn ThốiBelum ada peringkat
- Sordaria Lab ReportDokumen9 halamanSordaria Lab Reportapi-217409009Belum ada peringkat
- Parts of A Flower and Their FunctionsDokumen4 halamanParts of A Flower and Their FunctionsDaisuke InoueBelum ada peringkat
- Spring Syllable CardsDokumen12 halamanSpring Syllable CardsЖанна ПищикBelum ada peringkat
- Walker County Lake Birding ChecklistDokumen1 halamanWalker County Lake Birding ChecklistHap WalkerBelum ada peringkat
- 7.4 HomeostasisDokumen19 halaman7.4 HomeostasisJunkBelum ada peringkat
- Skeletal SystemDokumen5 halamanSkeletal SystemArra BeatrizBelum ada peringkat
- Chwache Breed of NepalDokumen11 halamanChwache Breed of NepalRoshan GiriBelum ada peringkat
- Peixes - FishDokumen8 halamanPeixes - FishlmviterboBelum ada peringkat
- Cementum PDFDokumen12 halamanCementum PDFHafra AminiBelum ada peringkat
- Frogdissectionpre Lab SiaDokumen2 halamanFrogdissectionpre Lab Siaapi-347123148Belum ada peringkat
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionDari EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionPenilaian: 4 dari 5 bintang4/5 (812)
- The Ugly History of Beautiful Things: Essays on Desire and ConsumptionDari EverandThe Ugly History of Beautiful Things: Essays on Desire and ConsumptionPenilaian: 4 dari 5 bintang4/5 (7)
- Exotic No More: Anthropology for the Contemporary WorldDari EverandExotic No More: Anthropology for the Contemporary WorldBelum ada peringkat
- Talking to the Enemy: Faith, Brotherhood, and the (Un)Making of TerroristsDari EverandTalking to the Enemy: Faith, Brotherhood, and the (Un)Making of TerroristsPenilaian: 4 dari 5 bintang4/5 (17)
- The Best Strangers in the World: Stories from a Life Spent ListeningDari EverandThe Best Strangers in the World: Stories from a Life Spent ListeningPenilaian: 4.5 dari 5 bintang4.5/5 (31)
- First Steps: How Upright Walking Made Us HumanDari EverandFirst Steps: How Upright Walking Made Us HumanPenilaian: 4 dari 5 bintang4/5 (17)
- Play: How it Shapes the Brain, Opens the Imagination, and Invigorates the SoulDari EverandPlay: How it Shapes the Brain, Opens the Imagination, and Invigorates the SoulPenilaian: 4 dari 5 bintang4/5 (96)
- The Web of Meaning: Integrating Science and Traditional Wisdom to Find our Place in the UniverseDari EverandThe Web of Meaning: Integrating Science and Traditional Wisdom to Find our Place in the UniversePenilaian: 5 dari 5 bintang5/5 (12)
- The Will to Change: Men, Masculinity, and LoveDari EverandThe Will to Change: Men, Masculinity, and LovePenilaian: 5 dari 5 bintang5/5 (384)
- Food of the Gods: The Search for the Original Tree of Knowledge: A Radical History of Plants, Drugs, and Human EvolutionDari EverandFood of the Gods: The Search for the Original Tree of Knowledge: A Radical History of Plants, Drugs, and Human EvolutionPenilaian: 4.5 dari 5 bintang4.5/5 (117)
- The Hundred Years' War on Palestine: A History of Settler Colonialism and Resistance, 1917–2017Dari EverandThe Hundred Years' War on Palestine: A History of Settler Colonialism and Resistance, 1917–2017Penilaian: 4 dari 5 bintang4/5 (63)
- They Thought They Were Free: The Germans, 1933-45Dari EverandThey Thought They Were Free: The Germans, 1933-45Penilaian: 4.5 dari 5 bintang4.5/5 (159)
- Civilized To Death: The Price of ProgressDari EverandCivilized To Death: The Price of ProgressPenilaian: 4.5 dari 5 bintang4.5/5 (215)
- The Immortality Key: The Secret History of the Religion with No NameDari EverandThe Immortality Key: The Secret History of the Religion with No NamePenilaian: 5 dari 5 bintang5/5 (3)
- Hunt, Gather, Parent: What Ancient Cultures Can Teach Us About the Lost Art of Raising Happy, Helpful Little HumansDari EverandHunt, Gather, Parent: What Ancient Cultures Can Teach Us About the Lost Art of Raising Happy, Helpful Little HumansPenilaian: 4.5 dari 5 bintang4.5/5 (176)
- Guns, Germs and Steel: The Fates of Human SocietiesDari EverandGuns, Germs and Steel: The Fates of Human SocietiesPenilaian: 4.5 dari 5 bintang4.5/5 (643)
- Sea People: The Puzzle of PolynesiaDari EverandSea People: The Puzzle of PolynesiaPenilaian: 4.5 dari 5 bintang4.5/5 (210)
- Generations: The Real Differences between Gen Z, Millennials, Gen X, Boomers, and Silents—and What They Mean for America's FutureDari EverandGenerations: The Real Differences between Gen Z, Millennials, Gen X, Boomers, and Silents—and What They Mean for America's FuturePenilaian: 4.5 dari 5 bintang4.5/5 (43)
- Mysterious Sightings In the SkyDari EverandMysterious Sightings In the SkyPenilaian: 4.5 dari 5 bintang4.5/5 (12)
- Nihongi: Chronicles of Japan from the Earliest of Times to A.D. 697Dari EverandNihongi: Chronicles of Japan from the Earliest of Times to A.D. 697Penilaian: 5 dari 5 bintang5/5 (2)
- Eight Bears: Mythic Past and Imperiled FutureDari EverandEight Bears: Mythic Past and Imperiled FuturePenilaian: 4.5 dari 5 bintang4.5/5 (5)
- Dark Matter of the Mind: The Culturally Articulated UnconsciousDari EverandDark Matter of the Mind: The Culturally Articulated UnconsciousPenilaian: 5 dari 5 bintang5/5 (1)
- The Wheel of Time: The Shamans of Mexico Their Thoughts about Life Death and the UniverseDari EverandThe Wheel of Time: The Shamans of Mexico Their Thoughts about Life Death and the UniversePenilaian: 4.5 dari 5 bintang4.5/5 (81)
- Anthro-Vision: A New Way to See in Business and LifeDari EverandAnthro-Vision: A New Way to See in Business and LifePenilaian: 4 dari 5 bintang4/5 (8)