Anda mungkin juga menyukai
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Aktivitas Promoter Â-Aktin Ikan Medaka Jepang (Oryzias Latipes) Pada Ikan Mas (Cyprinus Carpio)Dokumen8 halamanAktivitas Promoter Â-Aktin Ikan Medaka Jepang (Oryzias Latipes) Pada Ikan Mas (Cyprinus Carpio)TrafalgarSeieiBelum ada peringkat
- Biofloc Technology in Aquaculture: Beneficial Effects and Future ChallengesDokumen6 halamanBiofloc Technology in Aquaculture: Beneficial Effects and Future ChallengesTrafalgarSeieiBelum ada peringkat
- Comparison of Select Innate Immune Mechanisms of Fish and MammalsDokumen12 halamanComparison of Select Innate Immune Mechanisms of Fish and MammalsTrafalgarSeieiBelum ada peringkat
- SCientific WritingDokumen3 halamanSCientific Writingseila09Belum ada peringkat
- AquaDokumen297 halamanAquaTrafalgarSeieiBelum ada peringkat
- Comparisons of Chemical and Physical Properties of Catfish Oils Prepared From Different Extracting ProcessesDokumen7 halamanComparisons of Chemical and Physical Properties of Catfish Oils Prepared From Different Extracting ProcessesTrafalgarSeieiBelum ada peringkat
- Pakan Dengan Asam Lemak N-3Dokumen10 halamanPakan Dengan Asam Lemak N-3TrafalgarSeieiBelum ada peringkat
- Exotic Animal PracticeDokumen28 halamanExotic Animal PracticeTrafalgarSeieiBelum ada peringkat
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- XII Chemistry The P Block Elements1Dokumen50 halamanXII Chemistry The P Block Elements1vishalBelum ada peringkat
- October 2013 - Part 1 of 2Dokumen721 halamanOctober 2013 - Part 1 of 2SonjaWaltersBelum ada peringkat
- OIV-Methods Vol 1 en 2012Dokumen488 halamanOIV-Methods Vol 1 en 2012Violeta IP100% (1)
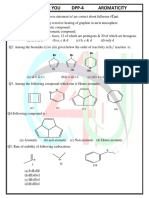
- Aromaticity DPP 4Dokumen4 halamanAromaticity DPP 4SubhadeepBelum ada peringkat
- METAL WORKS General CatalogueDokumen695 halamanMETAL WORKS General CatalogueSheila BowenBelum ada peringkat
- Review of Corrosion Inhibitors For Industrial Applications-1903Dokumen19 halamanReview of Corrosion Inhibitors For Industrial Applications-1903swoessner1100% (2)
- Effect of Natural Zeolite As Substrate Filler On The Properties of NBREPDM BlendDokumen8 halamanEffect of Natural Zeolite As Substrate Filler On The Properties of NBREPDM BlendhesBelum ada peringkat
- Material Information Data Sheet: Polyurethane - Flexible FoamDokumen8 halamanMaterial Information Data Sheet: Polyurethane - Flexible FoamPRASHANT SUDHAKAR SONAWANEBelum ada peringkat
- Lipid Digestion and AbsorptionDokumen37 halamanLipid Digestion and AbsorptionIMDCBiochem100% (1)
- GRANTED ABROAD (Based On E-Mail of IPU Dated (11/june/2021)Dokumen18 halamanGRANTED ABROAD (Based On E-Mail of IPU Dated (11/june/2021)Charowar HussainBelum ada peringkat
- Macromolecule Nutrient Lab ReportDokumen9 halamanMacromolecule Nutrient Lab ReportblakeBelum ada peringkat
- Science Class 10 Notes For Carbon and Its CompoundsDokumen4 halamanScience Class 10 Notes For Carbon and Its Compoundsmanal ahemadBelum ada peringkat
- (KBA BU HANUM) Senyawa Fenolik AlamDokumen134 halaman(KBA BU HANUM) Senyawa Fenolik AlamPoppyA.NamiraBelum ada peringkat
- Natural Products ChemistryDokumen26 halamanNatural Products Chemistrybharathi dasanBelum ada peringkat
- 11 Micro NutritionDokumen71 halaman11 Micro NutritionSumit RaiBelum ada peringkat
- STPP Hybrid Resin DesignDokumen4 halamanSTPP Hybrid Resin DesignPete WolanBelum ada peringkat
- Which One of Following Statements About The Fed and Fasting Metabolic States Is CorrectDokumen2 halamanWhich One of Following Statements About The Fed and Fasting Metabolic States Is Correctalvina damayantiBelum ada peringkat
- Controlled Delivery of Peptides and ProteinsDokumen19 halamanControlled Delivery of Peptides and ProteinsVanessya Oscar PermatasariBelum ada peringkat
- Penghitungan Obat Used Syringe PumpDokumen31 halamanPenghitungan Obat Used Syringe PumpFregiYuandiBelum ada peringkat
- Research Paper 2Dokumen45 halamanResearch Paper 2SushmaSahuBelum ada peringkat
- Moraxella Lacunata (Morax Axenfeld Bacillus or Haemophilus Duplex)Dokumen4 halamanMoraxella Lacunata (Morax Axenfeld Bacillus or Haemophilus Duplex)Mare5Der5Belum ada peringkat
- Chapter 2 Water ChemistryDokumen15 halamanChapter 2 Water ChemistryKathy Del CastilloBelum ada peringkat
- Drawback of Plastic Industry Cipet Final Report PDFDokumen32 halamanDrawback of Plastic Industry Cipet Final Report PDFSavi sharmaBelum ada peringkat
- 10.1.02 AOAC Official Method 985.22 Organochlorine and Organophosphorus Pesticide ResiduesDokumen0 halaman10.1.02 AOAC Official Method 985.22 Organochlorine and Organophosphorus Pesticide ResiduesTruong Hoai ThanhBelum ada peringkat
- Pigment DyeingDokumen28 halamanPigment DyeingAmad Mughal100% (1)
- Defix: Linacol MedicalDokumen2 halamanDefix: Linacol MedicalMohammed Al-YagoobBelum ada peringkat
- Preparation 19Dokumen3 halamanPreparation 19Kimberley Anne See100% (1)
- Chemicals' List and Mixing Protocol V9 - 06.05.2023Dokumen11 halamanChemicals' List and Mixing Protocol V9 - 06.05.2023Waqas AnjumBelum ada peringkat
- UNIT 1 Biology 3º EsoDokumen3 halamanUNIT 1 Biology 3º EsoHermi Rivas GarciaBelum ada peringkat
- Fertilizer and Pesticide Authority: Republic of The Philippines Department of AgricultureDokumen1 halamanFertilizer and Pesticide Authority: Republic of The Philippines Department of AgricultureJohn Henry NagaBelum ada peringkat