Anda mungkin juga menyukai
- Notas CerebroDokumen11 halamanNotas CerebroDaniie BayonaBelum ada peringkat
- Opioides en El Manejo Del Dolor.Dokumen12 halamanOpioides en El Manejo Del Dolor.jejoBelum ada peringkat
- Escalas NeurologicasDokumen6 halamanEscalas NeurologicasOkimus Angulo Vasquez100% (1)
- Fisiologia de La CocleaDokumen15 halamanFisiologia de La CocleaImim NozBelum ada peringkat
- Corteza CerebralDokumen11 halamanCorteza CerebralAndrea Fernández BlacuttBelum ada peringkat
- La Estructura Funcional Del Sistema NerviosoDokumen16 halamanLa Estructura Funcional Del Sistema Nerviosovaneza rebelde100% (2)
- Sindrome de Ramsay HuntDokumen1 halamanSindrome de Ramsay HuntRoberto MartinezBelum ada peringkat
- Funciones y circulación del LCR enDokumen4 halamanFunciones y circulación del LCR enJulio Jesus Sanchez Ordoñez100% (1)
- Cerebro Conducta....... Enfoques Que Describen La Relacion Cerebro-ConductaDokumen7 halamanCerebro Conducta....... Enfoques Que Describen La Relacion Cerebro-ConductaJohanna Ponce0% (1)
- NeurocisticercosisDokumen2 halamanNeurocisticercosisMaira Vargas OdicioBelum ada peringkat
- Exploración Del Aparato UrinarioDokumen8 halamanExploración Del Aparato UrinarioAndrea MedinaBelum ada peringkat
- Ciclo menstrual y regulación hormonalDokumen44 halamanCiclo menstrual y regulación hormonalJefferson LopezBelum ada peringkat
- La TEORÍA DEL DESARROLLO PSICOSEXUAL CON SUS DISTINTAS ETAPASDokumen3 halamanLa TEORÍA DEL DESARROLLO PSICOSEXUAL CON SUS DISTINTAS ETAPASnosferatus999Belum ada peringkat
- Resumen Libro de TanatologíaDokumen7 halamanResumen Libro de TanatologíaBeatriz SilvaBelum ada peringkat
- Pares CranealesDokumen6 halamanPares Cranealescamila ardilaBelum ada peringkat
- Qué Es La Trompa de EustaquioDokumen4 halamanQué Es La Trompa de EustaquioYael GuzmanBelum ada peringkat
- Arcos y Ángulos de Movimientos ArticularesDokumen15 halamanArcos y Ángulos de Movimientos ArticularesRaul RojasBelum ada peringkat
- Funciones Mentales SuperioresDokumen21 halamanFunciones Mentales SuperioresCarola Reyes MoncadaBelum ada peringkat
- Evc UtesaDokumen47 halamanEvc Utesajeison mesaBelum ada peringkat
- ElectrolitosDokumen4 halamanElectrolitosKevin Riera0% (1)
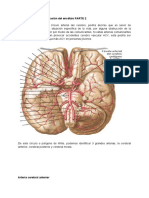
- Anatomía del sistema vascular cerebralDokumen5 halamanAnatomía del sistema vascular cerebralDanii Zamora MBelum ada peringkat
- HiponatremiaDokumen10 halamanHiponatremiaGabyflobeBelum ada peringkat
- Etapas Senectud y MuerteDokumen3 halamanEtapas Senectud y MuerteDiiana SuárezBelum ada peringkat
- Irrigacion CerebralDokumen14 halamanIrrigacion Cerebrallucy dragneelBelum ada peringkat
- Corrientes HumanistasDokumen30 halamanCorrientes HumanistasEvamary TG50% (2)
- Accidente Cerebro Vascular. PresentaciónDokumen65 halamanAccidente Cerebro Vascular. PresentaciónJoel Arrivillaga100% (2)
- Practica Agudeza Visual y AuditivaDokumen15 halamanPractica Agudeza Visual y AuditivaLaura JimenezBelum ada peringkat
- Desarrollo EmbrionarioDokumen16 halamanDesarrollo EmbrionarioDiayri DuarteBelum ada peringkat
- PSICOSISDokumen62 halamanPSICOSISbrenda olayoBelum ada peringkat
- Hipertension PulmonarDokumen33 halamanHipertension PulmonarAlejandro Escobar Huerta100% (1)
- Desordenes NeurologicosDokumen104 halamanDesordenes NeurologicosGlinny CastilloBelum ada peringkat
- Traumatismo Craneocefálico FISIOPATOLOGÍA Y MANEJODokumen38 halamanTraumatismo Craneocefálico FISIOPATOLOGÍA Y MANEJOLuis Daniel Hernández Torres100% (1)
- Aparato UrinarioDokumen34 halamanAparato UrinarioEstela Castro MontesBelum ada peringkat
- Psicofarmacología GeriátricaDokumen8 halamanPsicofarmacología GeriátricaBreinerd DazaBelum ada peringkat
- Caso Clinico NDokumen9 halamanCaso Clinico NCesarCumbrera0% (1)
- Exploracion NeurologicaDokumen216 halamanExploracion Neurologicalixal5910100% (1)
- Escalas de Valoración Funcional en El AncianoDokumen4 halamanEscalas de Valoración Funcional en El AncianoAdriana DíazBelum ada peringkat
- Cambios en La Piel Del Geronte PDFDokumen9 halamanCambios en La Piel Del Geronte PDFIvanovich NickBelum ada peringkat
- Enfermedad Vascular Cerebral (Evc)Dokumen22 halamanEnfermedad Vascular Cerebral (Evc)Ross Lee SüchtigkeitBelum ada peringkat
- Intoxicación Por OpioidesDokumen18 halamanIntoxicación Por OpioidesIrving BautistaBelum ada peringkat
- Citologia Lab ManualDokumen54 halamanCitologia Lab ManualQUIMICO CLINICO WILLIANS SANCHEZ100% (2)
- Protocolo Consumo de AlcoholDokumen41 halamanProtocolo Consumo de AlcoholKarinaKathryKat100% (1)
- Estados de la Conciencia: Sueño, Ritmos y TrastornosDokumen35 halamanEstados de la Conciencia: Sueño, Ritmos y TrastornosPatricia Leon100% (2)
- Aneurismas CerebralesDokumen21 halamanAneurismas CerebralesORUS GAMINGBelum ada peringkat
- Ptosis Palpebral Debilidad MuscularDokumen3 halamanPtosis Palpebral Debilidad MuscularJorge BecerraBelum ada peringkat
- Ética en CirugíaDokumen24 halamanÉtica en CirugíaMili Taipe RBelum ada peringkat
- 11 Enfermedades Crónicas No Transmisibles COSTA RICADokumen16 halaman11 Enfermedades Crónicas No Transmisibles COSTA RICAJorge Machado TencioBelum ada peringkat
- Anatomía del encéfalo y tronco encefálicoDokumen10 halamanAnatomía del encéfalo y tronco encefálicoMelissa MorilloBelum ada peringkat
- Sistema RespiratorioDokumen18 halamanSistema RespiratorioAlexandra Michelle100% (1)
- Intestino grueso: generalidades y segmentosDokumen113 halamanIntestino grueso: generalidades y segmentosJLuisCorderoC100% (1)
- Semiología GastrointestinalDokumen5 halamanSemiología GastrointestinalMary HeilanehBelum ada peringkat
- Capítulo 45 GuytonDokumen51 halamanCapítulo 45 GuytonBillcar Calderon88% (8)
- Cuaderno de TrabajoDokumen19 halamanCuaderno de TrabajoEduardoAlvearTorresBelum ada peringkat
- Organización Del Sistema NerviosoDokumen29 halamanOrganización Del Sistema NerviosoSindy Tatiana Valbuena CasallasBelum ada peringkat
- Deterioro CognitivoDokumen31 halamanDeterioro CognitivoGeraldineBelum ada peringkat
- Examen Físico Pares Craneales - Semiologia RaymundoDokumen35 halamanExamen Físico Pares Craneales - Semiologia RaymundoDiva Bruno OviedoBelum ada peringkat
- La Agresividad en La Conducta y Patología HumanaDokumen46 halamanLa Agresividad en La Conducta y Patología HumanaJosué CampilloBelum ada peringkat
- Páginas desdeAnatomia.y.Fisiologia - Saladin.6a.Edicion PDFDokumen2 halamanPáginas desdeAnatomia.y.Fisiologia - Saladin.6a.Edicion PDFdfgasdfasdf100% (1)
- Tema 9. Lenguaje, Mente y Cerebro 2Dokumen7 halamanTema 9. Lenguaje, Mente y Cerebro 2mariete018Belum ada peringkat
- Manual Lenguaje AdultosDokumen54 halamanManual Lenguaje AdultosDaniela Valdivia Flores100% (1)
- Bioelementos y BiomoleculasDokumen10 halamanBioelementos y BiomoleculasToita PonceBelum ada peringkat
- Informe Junio 2020 Lavado de ManosDokumen4 halamanInforme Junio 2020 Lavado de ManosToita Ponce100% (1)
- Informe Final de NutriciónDokumen9 halamanInforme Final de NutriciónToita PonceBelum ada peringkat
- Plan de Eventos AdversosDokumen5 halamanPlan de Eventos AdversosToita PonceBelum ada peringkat
- DISLIPIDEMIASDokumen24 halamanDISLIPIDEMIASToita PonceBelum ada peringkat
- Informe Sivan Mayo PS ChazoDokumen3 halamanInforme Sivan Mayo PS ChazoToita PonceBelum ada peringkat
- Informe Sivan Febrero PS ChazoDokumen4 halamanInforme Sivan Febrero PS ChazoToita PonceBelum ada peringkat
- Informe Sivan Enero PS ChazoDokumen4 halamanInforme Sivan Enero PS ChazoToita PonceBelum ada peringkat
- Informe Sivan Julio PS ChazoDokumen3 halamanInforme Sivan Julio PS ChazoToita PonceBelum ada peringkat
- Informe Sivan Agosto PS ChazoDokumen3 halamanInforme Sivan Agosto PS ChazoToita PonceBelum ada peringkat
- Informe Sivan Abril PS ChazoDokumen3 halamanInforme Sivan Abril PS ChazoToita PonceBelum ada peringkat
- FENILCETONURIADokumen7 halamanFENILCETONURIAToita PonceBelum ada peringkat
- LEUCEMIASDokumen12 halamanLEUCEMIASToita PonceBelum ada peringkat
- Fiebre AmarillaDokumen20 halamanFiebre AmarillaToita PonceBelum ada peringkat
- Informe FinalDokumen40 halamanInforme FinalToita PonceBelum ada peringkat
- Ponce 25Dokumen3 halamanPonce 25Toita PonceBelum ada peringkat
- Trastornos Vasculares Conferencia 1Dokumen42 halamanTrastornos Vasculares Conferencia 1Toita PonceBelum ada peringkat
- Atencion Del NiñoDokumen49 halamanAtencion Del NiñoToita PonceBelum ada peringkat
- Trastorno de La PersonalidadDokumen31 halamanTrastorno de La PersonalidadToita PonceBelum ada peringkat
- Canales Vaso Concepción y Vaso GobernadorDokumen26 halamanCanales Vaso Concepción y Vaso GobernadorToita PonceBelum ada peringkat
- Corrientes PsicológicasDokumen95 halamanCorrientes PsicológicasToita Ponce100% (1)
- LeucemiasDokumen30 halamanLeucemiasToita PonceBelum ada peringkat
- HIPERPOTASEMIADokumen20 halamanHIPERPOTASEMIAToita PonceBelum ada peringkat
- Cid TeoríaDokumen4 halamanCid TeoríaToita PonceBelum ada peringkat
- Cafe Maind Estudio de MercadoDokumen28 halamanCafe Maind Estudio de MercadoToita PonceBelum ada peringkat
- Artículo Científico PonceDokumen3 halamanArtículo Científico PonceToita PonceBelum ada peringkat
- Trastorno de La PersonalidadDokumen31 halamanTrastorno de La PersonalidadToita PonceBelum ada peringkat
- Asignacion de RecursosDokumen5 halamanAsignacion de RecursosToita PonceBelum ada peringkat
- Variables CualitativasDokumen25 halamanVariables CualitativasToita PonceBelum ada peringkat
- Ensayo ÉticaDokumen10 halamanEnsayo ÉticaToita PonceBelum ada peringkat
- Varela - El Concepto de Experiencia en AristotelesDokumen12 halamanVarela - El Concepto de Experiencia en AristotelesAlan GoodridgeBelum ada peringkat
- Dislexia en La Compresion Lectora - Seminario de InvestigacionDokumen33 halamanDislexia en La Compresion Lectora - Seminario de Investigacionyaquelin venturaBelum ada peringkat
- Tema 3 - Perspectivas Teóricas Del Desarrollo HumanoDokumen61 halamanTema 3 - Perspectivas Teóricas Del Desarrollo HumanoJose Ignacio Salcedo AlvarezBelum ada peringkat
- Evaluación Teorias Del AprendizajeDokumen3 halamanEvaluación Teorias Del AprendizajeJairo Ramos Aguirre100% (1)
- Descubriendo gustosDokumen2 halamanDescubriendo gustosCaroline QuezadaBelum ada peringkat
- Unidad 3, Modalidades de AprendizajeDokumen32 halamanUnidad 3, Modalidades de Aprendizajerhina diazBelum ada peringkat
- Ensayo PsicomotricidadDokumen8 halamanEnsayo PsicomotricidadnenacontadoraBelum ada peringkat
- Una Evaluación Formativa Enfocada en Competencias BuscaDokumen3 halamanUna Evaluación Formativa Enfocada en Competencias BuscaFrancisco Javier Fernandez HernandezBelum ada peringkat
- I. TEMARIO Resuelto IMPRESO.Dokumen79 halamanI. TEMARIO Resuelto IMPRESO.Nicole Cortés VidalBelum ada peringkat
- Mejora de atención en niños de 2 años con música clásicaDokumen156 halamanMejora de atención en niños de 2 años con música clásicaMariaBelum ada peringkat
- Las TIC y Su Influencia en Los Procesos de Enseñanza y AprendizajeDokumen13 halamanLas TIC y Su Influencia en Los Procesos de Enseñanza y AprendizajeDaniel GuapacheBelum ada peringkat
- Material de Lectura Unidad IIIDokumen23 halamanMaterial de Lectura Unidad IIIana lidia velazquez moralesBelum ada peringkat
- Trabaj Oral y Escrita - Margareth (Autoguardado)Dokumen9 halamanTrabaj Oral y Escrita - Margareth (Autoguardado)margareth racedoBelum ada peringkat
- Logoaudiometría en NiñosDokumen7 halamanLogoaudiometría en NiñosMariela CamposBelum ada peringkat
- Historia Del PsicodiagnósticoDokumen2 halamanHistoria Del PsicodiagnósticoBianca ElizaldeBelum ada peringkat
- Antología 2Dokumen230 halamanAntología 2HEMEROTECA0% (1)
- Trastornos neuropsicológicos: Alzheimer, autismo, dislexia y másDokumen5 halamanTrastornos neuropsicológicos: Alzheimer, autismo, dislexia y máswalter jesus navarro juarezBelum ada peringkat
- Documento Base Modulo 4Dokumen37 halamanDocumento Base Modulo 4asclepioBelum ada peringkat
- Sanando La Relación Que Tengo ConmigoDokumen74 halamanSanando La Relación Que Tengo ConmigoMariajose RiveraBelum ada peringkat
- Cartel de Las Capacidades y Sus Procesos CognitivosDokumen12 halamanCartel de Las Capacidades y Sus Procesos CognitivosPercy YactayoBelum ada peringkat
- Aprendizaje ExperiencialDokumen2 halamanAprendizaje ExperiencialHilda veronica SerranoBelum ada peringkat
- Tema 4. Estrategias de Apredizaje. Profesor Francisco Herrera Clavero.Dokumen10 halamanTema 4. Estrategias de Apredizaje. Profesor Francisco Herrera Clavero.Irene SfBelum ada peringkat
- Enfoque ComunicativoDokumen10 halamanEnfoque ComunicativoPortuon LopesBelum ada peringkat
- Etapas de La Vida HumanaDokumen4 halamanEtapas de La Vida HumanaIdkdeke RamiBelum ada peringkat
- WPPSI análisis FulanitoDokumen4 halamanWPPSI análisis FulanitoAna FerrufinoBelum ada peringkat
- Escribimos Rimas de AnimalesDokumen2 halamanEscribimos Rimas de AnimalesPATRICIA80% (5)
- Capítulo 6 Percepcion - (MP)Dokumen27 halamanCapítulo 6 Percepcion - (MP)NoeBelum ada peringkat
- PTU Lenguaje 2021Dokumen19 halamanPTU Lenguaje 2021Cronopia CortázarBelum ada peringkat
- Modelo de filtro rígido de BroadbentDokumen3 halamanModelo de filtro rígido de Broadbentvictor_dussan0% (1)