Anda mungkin juga menyukai
- Original Article: Endothelial Glycocalyx Damage Coincides With Microalbuminuria in Type 1 DiabetesDokumen6 halamanOriginal Article: Endothelial Glycocalyx Damage Coincides With Microalbuminuria in Type 1 DiabetesPandji 'Utuh'Belum ada peringkat
- Transforming Growth Factor (Type/3) Promotes The Addition of Chondroitin Sulfate Chains To The Cell Surface Proteoglycan (Syndecan) of Mouse Mammary EpitheliaDokumen10 halamanTransforming Growth Factor (Type/3) Promotes The Addition of Chondroitin Sulfate Chains To The Cell Surface Proteoglycan (Syndecan) of Mouse Mammary EpitheliaPandji 'Utuh'Belum ada peringkat
- Inflammation-And Ischemia-Induced Shedding of Venular GlycocalyxDokumen9 halamanInflammation-And Ischemia-Induced Shedding of Venular GlycocalyxPandji 'Utuh'Belum ada peringkat
- 14 Evan RubioDokumen10 halaman14 Evan RubioPandji 'Utuh'Belum ada peringkat
- Probing The Endothelial Cell Surface Layer (Glycocalyx) With Optical TweezersDokumen1 halamanProbing The Endothelial Cell Surface Layer (Glycocalyx) With Optical TweezersPandji 'Utuh'Belum ada peringkat
- Role of Reactive Oxygen Species in p53 Activation During Cisplatin-Induced Apoptosis of Rat Mesangial CellsDokumen7 halamanRole of Reactive Oxygen Species in p53 Activation During Cisplatin-Induced Apoptosis of Rat Mesangial CellsPandji 'Utuh'Belum ada peringkat
- The Diagnosis and Management of Chronic Cough: Ers Task ForceDokumen12 halamanThe Diagnosis and Management of Chronic Cough: Ers Task ForcePandji 'Utuh'Belum ada peringkat
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5795)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Cacophony BurnTheGroundDokumen32 halamanCacophony BurnTheGroundLeo HurtadoBelum ada peringkat
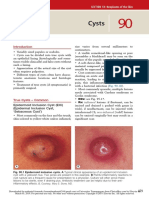
- CystsDokumen5 halamanCystsranindita maulyaBelum ada peringkat
- Ozone Unsung Hero by Navy Captain - Ajit VadakayilDokumen153 halamanOzone Unsung Hero by Navy Captain - Ajit Vadakayilsudhir shahBelum ada peringkat
- Examination of ElbowDokumen18 halamanExamination of Elbowharmohit singhBelum ada peringkat
- PhysiologyDokumen31 halamanPhysiologyraza20100% (1)
- Natural Science 6 PDFDokumen90 halamanNatural Science 6 PDFBeatriz Garcia67% (3)
- Autism Spectre DisorderDokumen13 halamanAutism Spectre DisorderJesus Hurtado100% (2)
- Drug Related Problem (DRP)Dokumen10 halamanDrug Related Problem (DRP)ERONADIAULFAH SUGITO50% (2)
- Aromatherapy For Professionals ENGDokumen593 halamanAromatherapy For Professionals ENGcamelia100% (3)
- Non - Food Meher Final Report (2023)Dokumen87 halamanNon - Food Meher Final Report (2023)atakltigebertsadikBelum ada peringkat
- Requests For MED3 Fit Note': Covid-19 Guidance For PracticesDokumen2 halamanRequests For MED3 Fit Note': Covid-19 Guidance For Practicesplayjake18Belum ada peringkat
- Documents 34-51Dokumen3 halamanDocuments 34-51api-575141270Belum ada peringkat
- HR 2 FormDokumen8 halamanHR 2 Formrkpatel40Belum ada peringkat
- AyyDokumen187 halamanAyyZoé WolfesBelum ada peringkat
- Intra-Arterial Catheterization For Invasive Monitoring: Indications, Insertion Techniques, and Interpretation - UpToDateDokumen40 halamanIntra-Arterial Catheterization For Invasive Monitoring: Indications, Insertion Techniques, and Interpretation - UpToDatejuanpbagurBelum ada peringkat
- Nursing Process Fon Chap 1Dokumen20 halamanNursing Process Fon Chap 1Saqlain M.Belum ada peringkat
- Rheumatic Fever PDFDokumen3 halamanRheumatic Fever PDFDoc JT BylnBelum ada peringkat
- Dermoscopy ReviewDokumen28 halamanDermoscopy ReviewMarwa RagabBelum ada peringkat
- Daily RevisionDokumen13 halamanDaily RevisionHanime hubBelum ada peringkat
- US - Inggris 2017Dokumen5 halamanUS - Inggris 2017SmabaitulhikmahBelum ada peringkat
- Introduction of The Topic: Nano-BiotechnologyDokumen7 halamanIntroduction of The Topic: Nano-BiotechnologyAbhishek YadavBelum ada peringkat
- Kjadjk BkjbasjdDokumen23 halamanKjadjk BkjbasjdzaenalBelum ada peringkat
- Heart Disease and Cancer: CirculationDokumen4 halamanHeart Disease and Cancer: CirculationDewi AyuBelum ada peringkat
- Michael H. Antoni, Roselyn Smith-Stress Management Intervention For Women With Breast Cancer-Amer Psychological Assn (2003)Dokumen222 halamanMichael H. Antoni, Roselyn Smith-Stress Management Intervention For Women With Breast Cancer-Amer Psychological Assn (2003)KmoespinoBelum ada peringkat
- Structural Organisation in Animals - IIDokumen70 halamanStructural Organisation in Animals - IIMay HarukaBelum ada peringkat
- English Composition II 1Dokumen11 halamanEnglish Composition II 1api-548473407Belum ada peringkat
- Bazaar DrugsDokumen104 halamanBazaar DrugsKartik Vashishta100% (1)
- Bril (2002) - Validation of The Toronto Clinical Scoring System For Diabetic PolyneuropathyDokumen5 halamanBril (2002) - Validation of The Toronto Clinical Scoring System For Diabetic PolyneuropathyRicky WatariBelum ada peringkat
- Vertex Announces Positive Day 90 Data For The First Patient in The Phase 1 - 2 Clinical Trial Dosed With VX-880, A Novel Investigational Stem Cell-Derived Therapy For The Treatment of Type 1 DiabetesDokumen3 halamanVertex Announces Positive Day 90 Data For The First Patient in The Phase 1 - 2 Clinical Trial Dosed With VX-880, A Novel Investigational Stem Cell-Derived Therapy For The Treatment of Type 1 Diabetesዘረአዳም ዘመንቆረርBelum ada peringkat
- Design Theory: Boo Virk Simon Andrews Boo - Virk@babraham - Ac.uk Simon - Andrews@babraham - Ac.ukDokumen33 halamanDesign Theory: Boo Virk Simon Andrews Boo - Virk@babraham - Ac.uk Simon - Andrews@babraham - Ac.ukuzma munirBelum ada peringkat