Anda mungkin juga menyukai
- Vitamin D and Fracture Risk in Early ChildhoodDokumen8 halamanVitamin D and Fracture Risk in Early ChildhoodpebinscribdBelum ada peringkat
- Lampiran JurnalDokumen3 halamanLampiran JurnalpebinscribdBelum ada peringkat
- Lion Air Eticket Itinerary / ReceiptDokumen2 halamanLion Air Eticket Itinerary / ReceiptpebinscribdBelum ada peringkat
- Cardiac ArrhytmiasDokumen4 halamanCardiac ArrhytmiaspebinscribdBelum ada peringkat
- Daftar Pustaka Bab IVDokumen1 halamanDaftar Pustaka Bab IVpebinscribdBelum ada peringkat
- MaternalandFetal Physiology - 111532Dokumen3 halamanMaternalandFetal Physiology - 111532viannikkkyBelum ada peringkat
- Accounting topics from introduction to strategic investment decisionsDokumen4 halamanAccounting topics from introduction to strategic investment decisionspebinscribdBelum ada peringkat
- Dapus BAB IIDokumen2 halamanDapus BAB IIpebinscribdBelum ada peringkat
- Lampiran JurnalDokumen3 halamanLampiran JurnalpebinscribdBelum ada peringkat
- Consumer BehaviorDokumen11 halamanConsumer BehaviorpebinscribdBelum ada peringkat
- Lanjutan Data Pasien DMDokumen1 halamanLanjutan Data Pasien DMpebinscribdBelum ada peringkat
- Microeconomics Concepts ExplainedDokumen14 halamanMicroeconomics Concepts ExplainedpebinscribdBelum ada peringkat
- J Neurol Neurosurg Psychiatry 2016 Noyce JNNP 2015 311890Dokumen9 halamanJ Neurol Neurosurg Psychiatry 2016 Noyce JNNP 2015 311890pebinscribdBelum ada peringkat
- Alrubaiee PDFDokumen25 halamanAlrubaiee PDFpebinscribdBelum ada peringkat
- 3 KonjungtivaDokumen42 halaman3 KonjungtivapebinscribdBelum ada peringkat
- J Neurol Neurosurg Psychiatry 2016 Dlouhy 402 13Dokumen13 halamanJ Neurol Neurosurg Psychiatry 2016 Dlouhy 402 13pebinscribdBelum ada peringkat
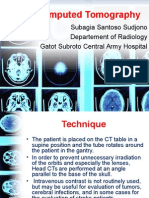
- Head Computed Tomography: Subagia Santoso Sudjono Departement of Radiology Gatot Subroto Central Army HospitalDokumen81 halamanHead Computed Tomography: Subagia Santoso Sudjono Departement of Radiology Gatot Subroto Central Army HospitalpebinscribdBelum ada peringkat
- J Neurol Neurosurg Psychiatry 2016 Gonçalves JNNP 2016 313458Dokumen7 halamanJ Neurol Neurosurg Psychiatry 2016 Gonçalves JNNP 2016 313458pebinscribdBelum ada peringkat
- Case Report: Tubocutaneous FistulaDokumen2 halamanCase Report: Tubocutaneous FistulapebinscribdBelum ada peringkat
- 2014 Article 369Dokumen6 halaman2014 Article 369pebinscribdBelum ada peringkat
- Guidelines Ambulatory Anesthesia SurgeryDokumen2 halamanGuidelines Ambulatory Anesthesia SurgerypebinscribdBelum ada peringkat
- Irdr 4 105Dokumen3 halamanIrdr 4 105pebinscribdBelum ada peringkat
- Paediatric Guidelines EUADokumen47 halamanPaediatric Guidelines EUApebinscribdBelum ada peringkat
- Irdr 4 105Dokumen3 halamanIrdr 4 105pebinscribdBelum ada peringkat
- ASA Status FisikDokumen1 halamanASA Status FisikpebinscribdBelum ada peringkat
- The Therapeutic Use of Analgesics in Patients With Liver CirrhosisDokumen7 halamanThe Therapeutic Use of Analgesics in Patients With Liver CirrhosispebinscribdBelum ada peringkat
- 2014 Article 369Dokumen6 halaman2014 Article 369pebinscribdBelum ada peringkat
- Zooterapia JornalEthnobiologyDokumen43 halamanZooterapia JornalEthnobiologyEdilene Coffaci de LimaBelum ada peringkat
- Kalaweit gibbon conservation programmeDokumen3 halamanKalaweit gibbon conservation programmepebinscribdBelum ada peringkat
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (119)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Vete 4305 - Group 2 Case Study - Cat With DiabetesDokumen19 halamanVete 4305 - Group 2 Case Study - Cat With Diabetesapi-242607157Belum ada peringkat
- Ramadan Health GuideDokumen30 halamanRamadan Health GuideISLAMICULTURE100% (2)
- Modifiable Stroke Risk FactorsDokumen2 halamanModifiable Stroke Risk FactorsPricilia GunawanBelum ada peringkat
- The Secrets of LongevityDokumen11 halamanThe Secrets of Longevitycarly183100% (10)
- How Australia identifies and addresses key health issuesDokumen28 halamanHow Australia identifies and addresses key health issuesCaleb FellowesBelum ada peringkat
- Epidemilogy Measurment MethodsDokumen100 halamanEpidemilogy Measurment MethodsKailash NagarBelum ada peringkat
- Case PresentationDokumen6 halamanCase PresentationYrrem UbaganBelum ada peringkat
- Closing The Quality Gap-A Critical Analysis of Quality Improvement StrategiesDokumen206 halamanClosing The Quality Gap-A Critical Analysis of Quality Improvement Strategiesturbina155Belum ada peringkat
- Hypertension: Dr. Lucia Mazur-Nicorici Md. PHDDokumen34 halamanHypertension: Dr. Lucia Mazur-Nicorici Md. PHDValerianBîcosBelum ada peringkat
- Specility DepartmentDokumen140 halamanSpecility DepartmentAjay SakarayBelum ada peringkat
- Mhra Annexure 10Dokumen40 halamanMhra Annexure 10Aqeel AhmedBelum ada peringkat
- Diabetes Types and Symptoms ExplainedDokumen77 halamanDiabetes Types and Symptoms ExplainedAshok100% (5)
- Type 2 Diabetes Menu Plan For Prevention and Management - FemaleDokumen9 halamanType 2 Diabetes Menu Plan For Prevention and Management - FemaleLuwani Lino100% (1)
- FBA Untuk Obat DMDokumen17 halamanFBA Untuk Obat DMKhoirunnisa MuslimawatiBelum ada peringkat
- Ladisch and Kohlmann 1992 - Recombinant Human InsulinDokumen10 halamanLadisch and Kohlmann 1992 - Recombinant Human InsulinZooey HeBelum ada peringkat
- Blood tests help diagnose diseasesDokumen10 halamanBlood tests help diagnose diseasesMay Mawzi WahabBelum ada peringkat
- English Nursing 2Dokumen22 halamanEnglish Nursing 2Anonymous fwaG1cNudUBelum ada peringkat
- Diabetes Mellitus in Pregnancy - Screening and DiagnosisDokumen7 halamanDiabetes Mellitus in Pregnancy - Screening and DiagnosisLuis Enrique Moscoso OrtizBelum ada peringkat
- Weaknes S/ Fatigue Polyphag IaDokumen5 halamanWeaknes S/ Fatigue Polyphag IaEwert Hesketh Nillama PaquinganBelum ada peringkat
- Accepted Paper in 3rd Medical ConferenceDokumen2 halamanAccepted Paper in 3rd Medical ConferenceMoha Mohammed100% (2)
- Deteksi Dini Kegawatdaruratan Maternal Dan Neonatal (RS Persada Medika Cikampek) PEBDokumen10 halamanDeteksi Dini Kegawatdaruratan Maternal Dan Neonatal (RS Persada Medika Cikampek) PEBindra pradjaBelum ada peringkat
- 1000 - 1500 AbstractBook - BookOfAbstractsDokumen500 halaman1000 - 1500 AbstractBook - BookOfAbstractsHasan Ibrahim KozanBelum ada peringkat
- Ashish Gajera B. Pharm. Project Roll No 14Dokumen116 halamanAshish Gajera B. Pharm. Project Roll No 14Ashish Gajera100% (4)
- Vildagliptin: The Evidence For Its Place in The Treatment of Type 2 Diabetes MellitusDokumen18 halamanVildagliptin: The Evidence For Its Place in The Treatment of Type 2 Diabetes Mellitusp k agarwalBelum ada peringkat
- Role of The Nurse in Stroke RehabilitationDokumen2 halamanRole of The Nurse in Stroke RehabilitationAnonymous mTqyS6Belum ada peringkat
- Healthy Eating Plate Guide to Balanced NutritionDokumen4 halamanHealthy Eating Plate Guide to Balanced Nutritionalexkanno100% (1)
- Other Health Impairment Fact SheetDokumen8 halamanOther Health Impairment Fact SheetNational Dissemination Center for Children with DisabilitiesBelum ada peringkat
- The Pathogenesis of The Diabetic Foot UlcerDokumen29 halamanThe Pathogenesis of The Diabetic Foot UlcernyuwwchocolavaBelum ada peringkat
- Austin Journal of Nutrition and Food SciencesDokumen7 halamanAustin Journal of Nutrition and Food SciencesAustin Publishing GroupBelum ada peringkat
- Family Medicine Exam Manual 2011Dokumen72 halamanFamily Medicine Exam Manual 2011SultanAlam100% (1)