Anda mungkin juga menyukai
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- Restu Dwi Apri Asih 14320064Dokumen2 halamanRestu Dwi Apri Asih 14320064Ahmad HabibiBelum ada peringkat
- Ahmad Fuad Hasan 14320052Dokumen5 halamanAhmad Fuad Hasan 14320052Ahmad HabibiBelum ada peringkat
- Teguh Widodo. 14320041Dokumen7 halamanTeguh Widodo. 14320041Ahmad HabibiBelum ada peringkat
- Ahmad Nugraha Tugastiyanto. 14320042Dokumen5 halamanAhmad Nugraha Tugastiyanto. 14320042Ahmad HabibiBelum ada peringkat
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (119)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- AntibioticsDokumen122 halamanAntibioticsPiyush Verma100% (2)
- Math 1030 Magnets and Pain ReliefDokumen6 halamanMath 1030 Magnets and Pain Reliefapi-444173009100% (1)
- Bolus Feeding in Adults A Practical GuideDokumen18 halamanBolus Feeding in Adults A Practical Guideshrutee babraBelum ada peringkat
- Prenatal Diagnosis PDFDokumen747 halamanPrenatal Diagnosis PDFFlavius Yo0% (1)
- Myocardial InfarctionDokumen4 halamanMyocardial InfarctionLetecia MooreBelum ada peringkat
- Assessment Form CPDokumen13 halamanAssessment Form CPvmsvino100% (2)
- Refresher Course: Preboard ExaminationDokumen6 halamanRefresher Course: Preboard ExaminationJastine SabornidoBelum ada peringkat
- Osimertinib or Platinum-Pemetrexed in EGFR T790M-Positive Lung CancerDokumen12 halamanOsimertinib or Platinum-Pemetrexed in EGFR T790M-Positive Lung CancerAdriana VladutuBelum ada peringkat
- Psychoanalisis and PaediatricsDokumen264 halamanPsychoanalisis and PaediatricsBlackKBelum ada peringkat
- Audiologists diagnose and treat hearing and balance issuesDokumen5 halamanAudiologists diagnose and treat hearing and balance issuesm afiq fahmiBelum ada peringkat
- Retinoblastoma - EyeWikiDokumen11 halamanRetinoblastoma - EyeWikimay171989Belum ada peringkat
- Drugs Acting On The Endocrine System Compiled WRDokumen74 halamanDrugs Acting On The Endocrine System Compiled WREyz PabloBelum ada peringkat
- Holistic Brain Injury Program for ChildrenDokumen12 halamanHolistic Brain Injury Program for ChildrenCatherineBelum ada peringkat
- NCP AppendectomyDokumen6 halamanNCP Appendectomymahmoud fuqahaBelum ada peringkat
- Reinforcement in The Classroom Improves Student Motivation and PerformanceDokumen4 halamanReinforcement in The Classroom Improves Student Motivation and PerformancePassmore DubeBelum ada peringkat
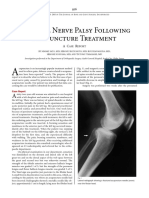
- Peroneal Nerve Palsy Following Acp TreatmentDokumen3 halamanPeroneal Nerve Palsy Following Acp TreatmentLydwiNa JcBelum ada peringkat
- Adrenergic Drugs Classification and UsesDokumen2 halamanAdrenergic Drugs Classification and UsesSunshine_Bacla_4275Belum ada peringkat
- Manual of Common Bedside Surgical Procedures-CHAPTER 9 Orthopedic ProcedureDokumen32 halamanManual of Common Bedside Surgical Procedures-CHAPTER 9 Orthopedic ProcedureCosti TanaseBelum ada peringkat
- Student Referral FormDokumen1 halamanStudent Referral FormMa Cyril Dawn MuncadaBelum ada peringkat
- MRI Safety Screening Form - Amended - 30NOV2016Dokumen1 halamanMRI Safety Screening Form - Amended - 30NOV2016FC LOONBelum ada peringkat
- Segmental Thoracic SpinalDokumen24 halamanSegmental Thoracic SpinalsoeBelum ada peringkat
- Adazet 6fig LN TCMWTDokumen13 halamanAdazet 6fig LN TCMWTSwagath NBelum ada peringkat
- Coughlin Community Health Paper IntroDokumen3 halamanCoughlin Community Health Paper Introapi-283315953Belum ada peringkat
- MCQ Haemato (DivC)Dokumen6 halamanMCQ Haemato (DivC)Muhammad Nazif0% (2)
- Nursing Home Investigation Tips Cheat SheetDokumen2 halamanNursing Home Investigation Tips Cheat SheetFreshyAndMessyBelum ada peringkat
- ID Hubungan Motivasi Perawat Dengan KepatuhDokumen10 halamanID Hubungan Motivasi Perawat Dengan KepatuhalfanBelum ada peringkat
- 5 Stages of Counseling ProsesDokumen16 halaman5 Stages of Counseling ProsesAraby Alfandani100% (1)
- Drug Abuse (Social Issue)Dokumen2 halamanDrug Abuse (Social Issue)IslamicGeneralKnowleBelum ada peringkat
- J. Bruce Bayne HIPPA FormDokumen5 halamanJ. Bruce Bayne HIPPA FormWilliam BayneBelum ada peringkat
- Source: Operating Room Technique by Nancymarie PhillipsDokumen1 halamanSource: Operating Room Technique by Nancymarie Phillipsnoreen ellieBelum ada peringkat