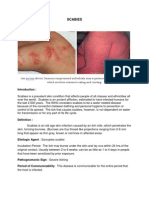
Anda mungkin juga menyukai
- All You Need to Know About Neck Threadworms and Your Itchy HorseDari EverandAll You Need to Know About Neck Threadworms and Your Itchy HorseBelum ada peringkat
- Common Mites (Arachnids) of Rabbits Common Mites (Arachnids) of Rabbits and Their Treatment and Their TreatmentDokumen8 halamanCommon Mites (Arachnids) of Rabbits Common Mites (Arachnids) of Rabbits and Their Treatment and Their TreatmentMustafarBelum ada peringkat
- Kelinci Notoedres Cati 1Dokumen16 halamanKelinci Notoedres Cati 1Nefertiti Rahmania IlmiBelum ada peringkat
- Bur, 2012Dokumen16 halamanBur, 2012Jessica VanyaBelum ada peringkat
- Ringworm Infection in Dogs and Cats: In: Recent Advances in Canine Infectious Diseases, L. Carmichael (Ed.)Dokumen7 halamanRingworm Infection in Dogs and Cats: In: Recent Advances in Canine Infectious Diseases, L. Carmichael (Ed.)Auni IbrahimBelum ada peringkat
- Insects and Allergic Reactions FixDokumen10 halamanInsects and Allergic Reactions FixFenty SuwidakBelum ada peringkat
- Scabies SRU Rev 06Dokumen31 halamanScabies SRU Rev 06Dwika HerdykiawanBelum ada peringkat
- Insects and Allergic Reactions: ) Department of Parasitology Faculty of Medicine, Trisakti UniversityDokumen7 halamanInsects and Allergic Reactions: ) Department of Parasitology Faculty of Medicine, Trisakti UniversityFenty SuwidakBelum ada peringkat
- Rabbit Skin Parasites 2002Dokumen9 halamanRabbit Skin Parasites 2002CabrasDeVillaraureBelum ada peringkat
- Demodicosis or Demodectic Mange Is A Disease That Affects The Cattle and IsDokumen3 halamanDemodicosis or Demodectic Mange Is A Disease That Affects The Cattle and IsElia JacobBelum ada peringkat
- Standardised Nomenclature of Animal Parasitic Diseases (Snopad)Dokumen67 halamanStandardised Nomenclature of Animal Parasitic Diseases (Snopad)Pwaveno BamaiyiBelum ada peringkat
- Fungi of Medical ImportanceDokumen11 halamanFungi of Medical ImportanceRi FitrianaBelum ada peringkat
- Infestation: ScabiesDokumen16 halamanInfestation: ScabiesBrix ValdrizBelum ada peringkat
- Dermatophytosis: LectureDokumen35 halamanDermatophytosis: LectureGideon MukalaziBelum ada peringkat
- Name: Joseph Olamide FaithDokumen5 halamanName: Joseph Olamide FaitholamideBelum ada peringkat
- 1) ArthropodsDokumen4 halaman1) Arthropodsyoupremium272727Belum ada peringkat
- Canine Anr Feline Dermatology & OtologyDokumen39 halamanCanine Anr Feline Dermatology & Otologycatita2389Belum ada peringkat
- Ectoparasites and Parasites On Human'S Skin/Body Surface: Parasitology DepartmentDokumen36 halamanEctoparasites and Parasites On Human'S Skin/Body Surface: Parasitology DepartmentmiaraissaBelum ada peringkat
- Ectoparasites and Parasites On Human'S Skin/Body Surface: Parasitology DepartmentDokumen36 halamanEctoparasites and Parasites On Human'S Skin/Body Surface: Parasitology DepartmentJimmy Fran IIBelum ada peringkat
- 3.11 MalariaDokumen26 halaman3.11 MalariaLince WijoyoBelum ada peringkat
- Pocket Pet DermatologyDokumen4 halamanPocket Pet DermatologyChris BernBelum ada peringkat
- ArthropodsDokumen42 halamanArthropodsKateBelum ada peringkat
- SCABIESDokumen14 halamanSCABIESNom Kumar Naik Rathod100% (1)
- Venomous AnimalsDokumen15 halamanVenomous AnimalsKaryl JoyBelum ada peringkat
- SOP For Staphylococcus AureusDokumen4 halamanSOP For Staphylococcus AureusDr. StormBelum ada peringkat
- (K6) Ectoparasites and Parasites On Human's Skinbody SurfaceDokumen38 halaman(K6) Ectoparasites and Parasites On Human's Skinbody SurfaceArifin MuhammadBelum ada peringkat
- Ectoparasites and Parasites On Human'S Skin/Body Surface: Parasitology DepartmentDokumen38 halamanEctoparasites and Parasites On Human'S Skin/Body Surface: Parasitology DepartmentArifin MuhammadBelum ada peringkat
- Ectoparasites and Endoparasites Affecting ReptilesDokumen8 halamanEctoparasites and Endoparasites Affecting ReptilesJohnLyndonLunnayBelum ada peringkat
- Dermatology TestsDokumen6 halamanDermatology TestsLouise Alysson OrtegaBelum ada peringkat
- Collecting and Preserving ArthropodsDokumen40 halamanCollecting and Preserving Arthropodsksambamuthi100% (6)
- Thelaziasis: Biology, Species Affected and Pathology (Conjunctivitis) : A ReviewDokumen2 halamanThelaziasis: Biology, Species Affected and Pathology (Conjunctivitis) : A ReviewAlla UddinBelum ada peringkat
- General Information About ScabiesDokumen16 halamanGeneral Information About ScabiesPramesti DarojahBelum ada peringkat
- Scabies in Indonesia: Causes, Transmission and SpeciesDokumen3 halamanScabies in Indonesia: Causes, Transmission and SpeciesMirlinda HartinBelum ada peringkat
- Treatment and Prevention of Pediculosis (Lice InfestationDokumen3 halamanTreatment and Prevention of Pediculosis (Lice InfestationAdrian MangahasBelum ada peringkat
- ScabiesDokumen8 halamanScabiesJessica Dumayas Dela RosaBelum ada peringkat
- Relieve Itchy Scabies with Proper TreatmentDokumen25 halamanRelieve Itchy Scabies with Proper TreatmentpmBelum ada peringkat
- Canine Atopic Dermatitis - A Practical ApproachDokumen15 halamanCanine Atopic Dermatitis - A Practical ApproachlirmvetBelum ada peringkat
- The Varroa mite parasite of honey beesDokumen6 halamanThe Varroa mite parasite of honey beesmanuel felipe villalbaBelum ada peringkat
- Papular Urticaria: SCRATCH Principles for Diagnosis and TreatmentDokumen2 halamanPapular Urticaria: SCRATCH Principles for Diagnosis and TreatmentanisaBelum ada peringkat
- Mycology Kopya 2 - 231122 - 203839Dokumen27 halamanMycology Kopya 2 - 231122 - 203839Çağrı İslam DemiralBelum ada peringkat
- Scabies: Scabies at A GlanceDokumen6 halamanScabies: Scabies at A GlanceseptaayuBelum ada peringkat
- Infection Control in HistopathDokumen13 halamanInfection Control in HistopathPA102294Belum ada peringkat
- Morphological Characteristics of TicksDokumen3 halamanMorphological Characteristics of TicksJane Xena RodriguezBelum ada peringkat
- Normal Flora of SkinDokumen14 halamanNormal Flora of SkinMedical StudentBelum ada peringkat
- Parasitismo en AvesDokumen23 halamanParasitismo en AvesCarolina De Los MonosBelum ada peringkat
- Vectors of Public Health ImportanceDokumen31 halamanVectors of Public Health ImportanceJoseph Stans Kasirye75% (4)
- Michigan Scabies Manual Provides GuidanceDokumen57 halamanMichigan Scabies Manual Provides Guidancevlada_cokoladaBelum ada peringkat
- Animal Science Lab 5 SheepDokumen5 halamanAnimal Science Lab 5 Sheepapi-355974802Belum ada peringkat
- Scabies (Calaustro Kyla)Dokumen22 halamanScabies (Calaustro Kyla)Carlojay IniegoBelum ada peringkat
- ARTICLE - Morgellons and Flying InsectsDokumen33 halamanARTICLE - Morgellons and Flying InsectsDaz93% (14)
- 3.02.01 AcarapisosisDokumen7 halaman3.02.01 Acarapisosismanuel felipe villalbaBelum ada peringkat
- Scabies NotesDokumen9 halamanScabies NotesAvi eBelum ada peringkat
- Sarcoptic Mange in A Dog: A Case Study: Original Research ArticleDokumen4 halamanSarcoptic Mange in A Dog: A Case Study: Original Research ArticlenadyasantikaBelum ada peringkat
- ScabiesDokumen2 halamanScabiesPRINTDESK by DanBelum ada peringkat
- Dermatophytes - Medical Research Institute - Alexandria UniversityDokumen129 halamanDermatophytes - Medical Research Institute - Alexandria Universityማላያላም ማላያላምBelum ada peringkat
- Economic ZoologyDokumen15 halamanEconomic ZoologySimamkeleBelum ada peringkat
- 1 s2.0 S2211753918301441 MainDokumen4 halaman1 s2.0 S2211753918301441 MainMateoBelum ada peringkat
- Arthropods PowerpointDokumen61 halamanArthropods PowerpointJaybee SarmientoBelum ada peringkat
- Prevalence of Dermatophytosis in Rabbits at SAQTVH, Chittagong, BangladeshDokumen6 halamanPrevalence of Dermatophytosis in Rabbits at SAQTVH, Chittagong, BangladeshNataly Belen Toribio VasquezBelum ada peringkat
- Invisible ItchesDokumen4 halamanInvisible ItchesGrace Saria MadridBelum ada peringkat
- Asme Sec. II Part A Ed. 2013 - Sa-451 To EndDokumen1.000 halamanAsme Sec. II Part A Ed. 2013 - Sa-451 To EndEnricoFurlan80% (5)
- Abpvc II Materialspart Dpropertiesmetric 140410201403 Phpapp01 PDFDokumen1.036 halamanAbpvc II Materialspart Dpropertiesmetric 140410201403 Phpapp01 PDFRoger RodriguesBelum ada peringkat
- Steel Castings Handbook Supplement 6 PDFDokumen101 halamanSteel Castings Handbook Supplement 6 PDFjperdigon9634Belum ada peringkat
- SCH, S9, High Alloy Data SheetsDokumen60 halamanSCH, S9, High Alloy Data SheetsMiki JaksicBelum ada peringkat
- S5-Steel Casting HandbookDokumen26 halamanS5-Steel Casting HandbookCharlie ChongBelum ada peringkat
- Steel Castings Handbook Supplement 6 PDFDokumen101 halamanSteel Castings Handbook Supplement 6 PDFjperdigon9634Belum ada peringkat
- HandBook Stuctural SteelDokumen267 halamanHandBook Stuctural SteelOnn Lajer100% (9)
- Process SafetyDokumen15 halamanProcess SafetyWormInchBelum ada peringkat
- KEYSTONE Butterfly and Rotary Valves SERIES 30,000Dokumen20 halamanKEYSTONE Butterfly and Rotary Valves SERIES 30,000WormInchBelum ada peringkat
- Ultrasonic Testing of Steel CastingDokumen38 halamanUltrasonic Testing of Steel CastingPARK, SANGYEOLBelum ada peringkat
- Steel Casting Handbook Supplement 8 - High Alloy Data Sheets - Corrosion SeriesDokumen93 halamanSteel Casting Handbook Supplement 8 - High Alloy Data Sheets - Corrosion SeriesjosemiguelzuBelum ada peringkat
- (Welding) s7Dokumen113 halaman(Welding) s7Popo YuppyBelum ada peringkat
- Steel Castings Harden AbilityDokumen31 halamanSteel Castings Harden AbilityAndrei State100% (1)
- Steel Castings Handbook PDFDokumen41 halamanSteel Castings Handbook PDFDave ImpresoBelum ada peringkat
- S5-Steel Casting HandbookDokumen26 halamanS5-Steel Casting HandbookCharlie ChongBelum ada peringkat
- Steel Casting Handbook Supplement 8 - High Alloy Data Sheets - Corrosion SeriesDokumen93 halamanSteel Casting Handbook Supplement 8 - High Alloy Data Sheets - Corrosion SeriesjosemiguelzuBelum ada peringkat
- Steel Casting Handbook Supplement 8 - High Alloy Data Sheets - Corrosion SeriesDokumen93 halamanSteel Casting Handbook Supplement 8 - High Alloy Data Sheets - Corrosion SeriesjosemiguelzuBelum ada peringkat
- HandBook Stuctural SteelDokumen267 halamanHandBook Stuctural SteelOnn Lajer100% (9)
- SCH, S9, High Alloy Data SheetsDokumen60 halamanSCH, S9, High Alloy Data SheetsMiki JaksicBelum ada peringkat
- Steel Casting HandbookDokumen74 halamanSteel Casting Handbooktang weng wai100% (2)
- Steel Castings Handbook Supplement 6 PDFDokumen101 halamanSteel Castings Handbook Supplement 6 PDFjperdigon9634Belum ada peringkat
- Suplement4 Steel Casting NadbookDokumen8 halamanSuplement4 Steel Casting NadbookDiego Lorenzo AparicioBelum ada peringkat
- Ultrasonic Testing of Steel CastingDokumen38 halamanUltrasonic Testing of Steel CastingPARK, SANGYEOLBelum ada peringkat
- (Welding) s7Dokumen113 halaman(Welding) s7Popo YuppyBelum ada peringkat
- Steel Castings Handbook PDFDokumen41 halamanSteel Castings Handbook PDFDave ImpresoBelum ada peringkat
- S 1Dokumen25 halamanS 1Grigoris PavlakisBelum ada peringkat
- Steel Castings Harden AbilityDokumen31 halamanSteel Castings Harden AbilityAndrei State100% (1)
- HandBook Stuctural SteelDokumen267 halamanHandBook Stuctural SteelOnn Lajer100% (9)
- S5-Steel Casting HandbookDokumen26 halamanS5-Steel Casting HandbookCharlie ChongBelum ada peringkat
- SCH, S9, High Alloy Data SheetsDokumen60 halamanSCH, S9, High Alloy Data SheetsMiki JaksicBelum ada peringkat
- Scabies - Epidemiology, Clinical Features, and Diagnosis - UpToDateDokumen61 halamanScabies - Epidemiology, Clinical Features, and Diagnosis - UpToDateJorge ArcentalesBelum ada peringkat
- ParasiticDokumen106 halamanParasitichelloaBelum ada peringkat
- 1 s2.0 S2214250923002007 MainDokumen2 halaman1 s2.0 S2214250923002007 MainsafitrinadiawahyuBelum ada peringkat
- Effectiveness of Madre de CacaoDokumen17 halamanEffectiveness of Madre de CacaoAthena Noceto82% (17)
- Itchy Infestation: The Causes and Treatment of ScabiesDokumen39 halamanItchy Infestation: The Causes and Treatment of ScabiesEx CalioseBelum ada peringkat
- Scabies NotesDokumen9 halamanScabies NotesAvi eBelum ada peringkat
- ScabiesDokumen38 halamanScabiesSiti Sabrina AtamiaBelum ada peringkat
- Scabies NCPDokumen6 halamanScabies NCPangeliejoy_1109100% (4)
- Scabicidal ResearchDokumen73 halamanScabicidal ResearchLovely Ann AyapanaBelum ada peringkat
- Dombing DiseaseDokumen5 halamanDombing Diseasemuhammad YunusBelum ada peringkat
- Fleas, Ticks, Lice, Mites, and Flies: Animal HealthDokumen8 halamanFleas, Ticks, Lice, Mites, and Flies: Animal HealthAwwal DayyabuBelum ada peringkat
- Entomology Item6,7Dokumen69 halamanEntomology Item6,7Barat NiloyBelum ada peringkat
- Jurnal Scabies 7Dokumen9 halamanJurnal Scabies 7Eric FullerBelum ada peringkat
- Skin ScrapeDokumen7 halamanSkin ScrapeShanmathiBelum ada peringkat
- إستبيانDokumen11 halamanإستبيانمرام محمد بخيت خيريBelum ada peringkat
- Medical EntomologyDokumen39 halamanMedical EntomologyRIZIQ TSBelum ada peringkat
- Scabies SRUDokumen20 halamanScabies SRUHesti Prihastuti DewiBelum ada peringkat
- Scabies PresentationDokumen15 halamanScabies Presentationrhimineecat71Belum ada peringkat
- 4.medical Entomology and Vector ControlDokumen68 halaman4.medical Entomology and Vector ControlMeutia Wilda100% (4)
- Dec 2010Dokumen94 halamanDec 2010Faiz DhmBelum ada peringkat
- Animal Diseases CollectionDokumen94 halamanAnimal Diseases CollectionNabin Kumar YadavBelum ada peringkat
- Entomology Lab VETM 2011 - 2021Dokumen13 halamanEntomology Lab VETM 2011 - 2021TheBoss 20Belum ada peringkat
- Scabies MGMT PP TDokumen31 halamanScabies MGMT PP TIman QaisarahBelum ada peringkat
- Scabies by DR Najeeb Memon PakistanDokumen39 halamanScabies by DR Najeeb Memon Pakistanmnajeeb807196Belum ada peringkat
- Sarcoptes Scabiei Mite Life CycleDokumen7 halamanSarcoptes Scabiei Mite Life CycleSiti Zainab Bani PurwanaBelum ada peringkat
- Module 3 HomeworkDokumen6 halamanModule 3 HomeworkCj LinceBelum ada peringkat
- Treatment of Crusted Scabies With Albendazole - A Case Report (EScholarship)Dokumen3 halamanTreatment of Crusted Scabies With Albendazole - A Case Report (EScholarship)pakemainmain100% (2)
- House FlyDokumen45 halamanHouse FlysanthiyasandyBelum ada peringkat
- Microbial Diseases of The Skin and Eyes (Microbiology Chapter 21)Dokumen82 halamanMicrobial Diseases of The Skin and Eyes (Microbiology Chapter 21)Katrina Isabel100% (1)