Anda mungkin juga menyukai
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- MP - Hardgainer GuideDokumen14 halamanMP - Hardgainer GuideAnthony Dinicolantonio100% (2)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Reiki Wings - Rinku PatelDokumen82 halamanReiki Wings - Rinku PatelOana Simionescu100% (3)
- 12-Fancy Birds RearingDokumen45 halaman12-Fancy Birds RearingAnonymous sBJu38BqBelum ada peringkat
- Samantha at Saratoga, Or, "Racin' After Fashion"Dokumen392 halamanSamantha at Saratoga, Or, "Racin' After Fashion"DanielBelum ada peringkat
- Yoga Basics For Men PDFDokumen47 halamanYoga Basics For Men PDFfludor100% (1)
- Common Medical AbbriviationDokumen13 halamanCommon Medical AbbriviationADAMU LAWALBelum ada peringkat
- Assessment of The Ear, Nose and ThroatDokumen40 halamanAssessment of The Ear, Nose and Throatsnickers_j100% (3)
- Anna Rozaliyani: Department of Parasitology-Faculty of Medicine, Universitas IndonesiaDokumen27 halamanAnna Rozaliyani: Department of Parasitology-Faculty of Medicine, Universitas Indonesiainka.elseBelum ada peringkat
- Gene Expression Regulation of Procaryotes & Viruses: Fera Ibrahim & T. Mirawati Sudiro Departemen Mikrobiologi FkuiDokumen80 halamanGene Expression Regulation of Procaryotes & Viruses: Fera Ibrahim & T. Mirawati Sudiro Departemen Mikrobiologi Fkuiinka.elseBelum ada peringkat
- Intro To The Function of Thyroid GlandDokumen35 halamanIntro To The Function of Thyroid Glandinka.elseBelum ada peringkat
- Intro To The Function of Thyroid GlandDokumen35 halamanIntro To The Function of Thyroid Glandinka.elseBelum ada peringkat
- Intro Tothe Function of Parathyroid Gland, ConciseDokumen34 halamanIntro Tothe Function of Parathyroid Gland, Conciseinka.elseBelum ada peringkat
- Introduction To The Endocrine Control On MetabolismDokumen79 halamanIntroduction To The Endocrine Control On Metabolisminka.elseBelum ada peringkat
- Thyroid and Antithyroid DrugsDokumen49 halamanThyroid and Antithyroid Drugsinka.elseBelum ada peringkat
- Effects of Bimanual Functional Practice Training On Functional Performance of Upper Extremity in Chronic Stroke SRJI Vol 2 Issue 3 Year 2013Dokumen10 halamanEffects of Bimanual Functional Practice Training On Functional Performance of Upper Extremity in Chronic Stroke SRJI Vol 2 Issue 3 Year 2013Dr. Krishna N. SharmaBelum ada peringkat
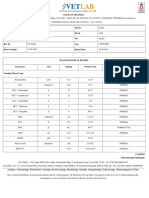
- Haematology, Biochemistry, Serology and Immunology, Microbiology, Cytology, Histopathology, Endocrinology, Ultrasonography & X-RayDokumen2 halamanHaematology, Biochemistry, Serology and Immunology, Microbiology, Cytology, Histopathology, Endocrinology, Ultrasonography & X-RayPankaj BeniwalBelum ada peringkat
- Hamster Biology HusbandryDokumen39 halamanHamster Biology HusbandryKulenović Zlatan100% (1)
- Sakool ProfileDokumen3 halamanSakool ProfileksakoolBelum ada peringkat
- Duchenne Muscular Dystrophy: A Cerebellar Disorder?: Shana E. Cyrulnik, Veronica J. HintonDokumen11 halamanDuchenne Muscular Dystrophy: A Cerebellar Disorder?: Shana E. Cyrulnik, Veronica J. HintonFajri MuhammadBelum ada peringkat
- GuppyDokumen2 halamanGuppymartynunez12345Belum ada peringkat
- Duck Rearing and ManagemntDokumen49 halamanDuck Rearing and Managemntkaranpraba1901Belum ada peringkat
- MaternityDokumen91 halamanMaternityAnonymous D8KswoBelum ada peringkat
- Cestodes 2020Dokumen10 halamanCestodes 2020CDBelum ada peringkat
- 2.biomekanik Pada Edentulus PenuhDokumen27 halaman2.biomekanik Pada Edentulus PenuhJesica Dwiasta Octaria NainggolanBelum ada peringkat
- Ielts 5.0 l1 R DoneDokumen14 halamanIelts 5.0 l1 R DoneMai TrangBelum ada peringkat
- Tissue TableDokumen4 halamanTissue TableMarcie BooneBelum ada peringkat
- Syphilis - The Great ImitatorDokumen4 halamanSyphilis - The Great ImitatorIJAERS JOURNALBelum ada peringkat
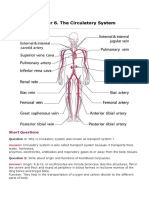
- Chapter 6. The Circulatory System: Short QuestionsDokumen31 halamanChapter 6. The Circulatory System: Short QuestionsParardha DharBelum ada peringkat
- Parasitology Case Presentation: Group 3Dokumen42 halamanParasitology Case Presentation: Group 3Christine Joy LegaspiBelum ada peringkat
- Double Crush SyndromeDokumen14 halamanDouble Crush SyndromeJosip SulenticBelum ada peringkat
- 24-06-2020 HMB EnglishDokumen20 halaman24-06-2020 HMB EnglishRakshith GowdaBelum ada peringkat
- Medical Parasitology: Human ParasitesDokumen30 halamanMedical Parasitology: Human ParasitesElycaBelum ada peringkat
- Urinary SystemDokumen31 halamanUrinary SystemAyro Business CenterBelum ada peringkat
- MCQSDokumen3 halamanMCQSShahzad RasoolBelum ada peringkat
- Tula at Reviewer For PrintDokumen14 halamanTula at Reviewer For PrintMariejho GensayaBelum ada peringkat
- Practice Prelim 3-KeyDokumen3 halamanPractice Prelim 3-KeyLauren PriscoBelum ada peringkat
- Nodal Stations of The AbdomenDokumen8 halamanNodal Stations of The Abdomenborst0% (1)