Anda mungkin juga menyukai
- Tmp72e2 TMPDokumen14 halamanTmp72e2 TMPFrontiersBelum ada peringkat
- The Pathophysiology of Motor Symptoms in Parkinson's DiseaseDokumen6 halamanThe Pathophysiology of Motor Symptoms in Parkinson's DiseasegracegozaliBelum ada peringkat
- Role of Preclinical Data in PDDokumen23 halamanRole of Preclinical Data in PDIrving ParraBelum ada peringkat
- Mtanpclbp FinalDokumen12 halamanMtanpclbp FinalSachin AbsBelum ada peringkat
- Biol Psychiatr 2014 Nov 76 (9) 678Dokumen3 halamanBiol Psychiatr 2014 Nov 76 (9) 678drclaudiofernandomedBelum ada peringkat
- Frontal Brain Activity and Cognitive Processing Speed in Multiple SclerosisDokumen8 halamanFrontal Brain Activity and Cognitive Processing Speed in Multiple SclerosisGeo MeBelum ada peringkat
- 10 1016@j Clinph 2019 09 021Dokumen37 halaman10 1016@j Clinph 2019 09 021Jihen FouratiBelum ada peringkat
- 5 - Aerobic Exercise Alters Brain Function and Structure in Parkinson's Disease A Randomized Controlled TrialDokumen14 halaman5 - Aerobic Exercise Alters Brain Function and Structure in Parkinson's Disease A Randomized Controlled TrialGabriel NevesBelum ada peringkat
- Stephan 2006 PlasticityDokumen11 halamanStephan 2006 PlasticityGiuli PanizzoloBelum ada peringkat
- Chiken 2014Dokumen9 halamanChiken 2014dgdgfgfBelum ada peringkat
- Brainsci 09 00005Dokumen8 halamanBrainsci 09 00005Javier Leonardo Rocha BeltranBelum ada peringkat
- Deep Brain Stimulation in 2018Dokumen11 halamanDeep Brain Stimulation in 2018Dhino Armand Quispe SánchezBelum ada peringkat
- Motor Priming in NeurorehabilitationDokumen15 halamanMotor Priming in NeurorehabilitationGhea Mae VillasBelum ada peringkat
- Ni Hms 749680Dokumen32 halamanNi Hms 749680Cristian GhiţăBelum ada peringkat
- Biology ProjectDokumen14 halamanBiology ProjectVittal Rao LBelum ada peringkat
- J NBD 2021 105372Dokumen10 halamanJ NBD 2021 105372bobBelum ada peringkat
- tmp9F14 TMPDokumen24 halamantmp9F14 TMPFrontiersBelum ada peringkat
- Neuro-Cardiac-guided TMSDokumen11 halamanNeuro-Cardiac-guided TMSRafael MoreiraBelum ada peringkat
- BOGGIO Tdcs Alcohol CravingDokumen6 halamanBOGGIO Tdcs Alcohol Cravingexcelencia5Belum ada peringkat
- Grosenick Neuron 2015Dokumen34 halamanGrosenick Neuron 2015AntonellaBelum ada peringkat
- Ambulation and Parkinson DiseaseDokumen22 halamanAmbulation and Parkinson DiseaseDeborah SalinasBelum ada peringkat
- Cognition and Mood in Parkinson's Disease in Subthalamic Nucleus Versus Globus Pallidus Interna Deep Brain Stimulation The COMPARE TrialDokumen10 halamanCognition and Mood in Parkinson's Disease in Subthalamic Nucleus Versus Globus Pallidus Interna Deep Brain Stimulation The COMPARE TrialRicardo GarciaBelum ada peringkat
- Neurostimulation For Stroke RehabilitationDokumen11 halamanNeurostimulation For Stroke Rehabilitationisos.mporei.vevaios.Belum ada peringkat
- Mindfullnes and CongnitionDokumen10 halamanMindfullnes and CongnitionCamila AmorimBelum ada peringkat
- EEG Biofeedback of Low Beta Band Components: Frequency-Specific Effects On Variables of Attention and Event-Related Brain PotentialsDokumen9 halamanEEG Biofeedback of Low Beta Band Components: Frequency-Specific Effects On Variables of Attention and Event-Related Brain PotentialsIshika SharmaBelum ada peringkat
- J Parkreldis 2015 09 028Dokumen4 halamanJ Parkreldis 2015 09 028T.U. Pai João da Caridade & Caboclo FlechasBelum ada peringkat
- Margarita Zachariou Et Al - Sensory Gating and Its Modulation by Cannabinoids: Electrophysiological, Computational and Mathematical AnalysisDokumen12 halamanMargarita Zachariou Et Al - Sensory Gating and Its Modulation by Cannabinoids: Electrophysiological, Computational and Mathematical AnalysisJmasnBelum ada peringkat
- NMDA Receptor Hypofunction Produces Concomitant Firing Rate Potentiation and Burst Activity Reduction in The Prefrontal CortexDokumen6 halamanNMDA Receptor Hypofunction Produces Concomitant Firing Rate Potentiation and Burst Activity Reduction in The Prefrontal CortexCortate15gBelum ada peringkat
- Delayed Plastic Responses To Anodal TDCS in Older Adults. Fujiyama Et Al 2014Dokumen9 halamanDelayed Plastic Responses To Anodal TDCS in Older Adults. Fujiyama Et Al 2014Saray Parra TorróBelum ada peringkat
- Model Responses For Additional SAQ'S PaperDokumen14 halamanModel Responses For Additional SAQ'S PaperYoyo YuBelum ada peringkat
- How Psychotherapy Changes The BrainDokumen5 halamanHow Psychotherapy Changes The BrainTomas SærmarkBelum ada peringkat
- Schulz Gerloff Hummel 2013 Non Invasive Brain Stimulation in Neurological DiseasesDokumen9 halamanSchulz Gerloff Hummel 2013 Non Invasive Brain Stimulation in Neurological DiseasesandreapalmapBelum ada peringkat
- Cholinergic Modulation of Skill Learning and PlasticityDokumen3 halamanCholinergic Modulation of Skill Learning and PlasticityPablo Arenas MancillaBelum ada peringkat
- BHX 055Dokumen13 halamanBHX 055Alfeku CabraBelum ada peringkat
- Irrelevant Stimulus Processing in ADHD: Catecholamine Dynamics and Attentional NetworksDokumen15 halamanIrrelevant Stimulus Processing in ADHD: Catecholamine Dynamics and Attentional NetworksMarie MamatBelum ada peringkat
- Default Mode Network and MeditationDokumen16 halamanDefault Mode Network and MeditationJustin StanleyBelum ada peringkat
- The Utility of Non-Invasive Brain Stimulation in RDokumen10 halamanThe Utility of Non-Invasive Brain Stimulation in RsanagaumiBelum ada peringkat
- Neurophysiological Bio Markers For Drug Development in Schizophrenia. Nat Rev Drug Discov 2008Dokumen16 halamanNeurophysiological Bio Markers For Drug Development in Schizophrenia. Nat Rev Drug Discov 2008Ignacio VarelaBelum ada peringkat
- Transcranial Magnetic Stimulation: Juan Jose Lo Pez-Ibor, Marı A Ine S Lo Pez-Ibor and Jose Ignacio PastranaDokumen7 halamanTranscranial Magnetic Stimulation: Juan Jose Lo Pez-Ibor, Marı A Ine S Lo Pez-Ibor and Jose Ignacio PastranaJ Ignacio PastranaBelum ada peringkat
- PMS, EEG, and Photic StimulationDokumen7 halamanPMS, EEG, and Photic StimulationmarinicamicaBelum ada peringkat
- Neuroplasticity and Non-Invasive Brain Stimulation in The Developing BrainDokumen33 halamanNeuroplasticity and Non-Invasive Brain Stimulation in The Developing BrainRebeca FloresBelum ada peringkat
- Improving Working Memory Exploring The Effect of Transcranial RandomDokumen6 halamanImproving Working Memory Exploring The Effect of Transcranial RandomDanish MujibBelum ada peringkat
- RTMS Gait Parkinsonism Kim2015Dokumen10 halamanRTMS Gait Parkinsonism Kim2015Eduardo Fernández DortaBelum ada peringkat
- How Electroconvulsive Therapy Works - Understanding The Neurobiological Mechanisms - PMCDokumen23 halamanHow Electroconvulsive Therapy Works - Understanding The Neurobiological Mechanisms - PMCSuraj DubeyBelum ada peringkat
- Neurobiology Paper FinalDokumen10 halamanNeurobiology Paper Finalapi-583318017Belum ada peringkat
- NIH Public Access: Author ManuscriptDokumen27 halamanNIH Public Access: Author ManuscriptVictor CarrenoBelum ada peringkat
- Jpts 27 673Dokumen4 halamanJpts 27 673Lazlo SecretBelum ada peringkat
- Pulsed Near Infrared Transcranial and Intranasal Photobiomodulation Significantly Modulates Neural Oscillations: A Pilot Exploratory StudyDokumen11 halamanPulsed Near Infrared Transcranial and Intranasal Photobiomodulation Significantly Modulates Neural Oscillations: A Pilot Exploratory StudyAndré RamosBelum ada peringkat
- Neuro FeedbackDokumen14 halamanNeuro FeedbackMar LindaBelum ada peringkat
- Placebo Article in ArchivesDokumen12 halamanPlacebo Article in ArchivesIsak Isakov100% (2)
- Percutaneous Tibial Nerve Stimulation (PTNS) Efficacy in The Treatment of Lower Urinary Tract Dysfunctions: A Systematic ReviewDokumen11 halamanPercutaneous Tibial Nerve Stimulation (PTNS) Efficacy in The Treatment of Lower Urinary Tract Dysfunctions: A Systematic ReviewMUHAMMMAD FAKHRIBelum ada peringkat
- The Influence of Dopamine On Spatial VisionDokumen7 halamanThe Influence of Dopamine On Spatial VisiontratrBelum ada peringkat
- #6479 Bliem B 2008Dokumen12 halaman#6479 Bliem B 2008marcu_ruxandraBelum ada peringkat
- Papel de La Microglía en La SinapsisDokumen13 halamanPapel de La Microglía en La SinapsisLorena de la FlorBelum ada peringkat
- TMP 13 F2Dokumen16 halamanTMP 13 F2FrontiersBelum ada peringkat
- Toxins: Central Effects of Botulinum Neurotoxin-Evidence From Human StudiesDokumen11 halamanToxins: Central Effects of Botulinum Neurotoxin-Evidence From Human StudiesTameemBelum ada peringkat
- Neuropsychological Rehabilitation, Neuroimaging and Neuropasticity A Clinical CommentaryDokumen11 halamanNeuropsychological Rehabilitation, Neuroimaging and Neuropasticity A Clinical CommentaryRossana Maciel Taboada GarzónBelum ada peringkat
- An Independent Components and Functional Connectivity Analysis of Resting State FMRI Data Points To Neural Network Dysregulation in Adult ADHDDokumen12 halamanAn Independent Components and Functional Connectivity Analysis of Resting State FMRI Data Points To Neural Network Dysregulation in Adult ADHDValentina ScorzoneBelum ada peringkat
- The Properties and Connections of Nerve Cells in Leech Ganglia Maintained in CultureDokumen19 halamanThe Properties and Connections of Nerve Cells in Leech Ganglia Maintained in CultureMaría Celeste Sánchez SugíaBelum ada peringkat
- Requirement of AQP4 For Antidepressive Efficiency of Fluoxetine: Implication in Adult Hippocampal NeurogenesisDokumen14 halamanRequirement of AQP4 For Antidepressive Efficiency of Fluoxetine: Implication in Adult Hippocampal NeurogenesisMaría Celeste Sánchez SugíaBelum ada peringkat
- Hodgkin Katz 1949Dokumen41 halamanHodgkin Katz 1949María Celeste Sánchez SugíaBelum ada peringkat
- Neurobiology of Learning and Memory: Nahid Majlessi, Samira Choopani, Tahereh Bozorgmehr, Zahra AziziDokumen7 halamanNeurobiology of Learning and Memory: Nahid Majlessi, Samira Choopani, Tahereh Bozorgmehr, Zahra AziziMaría Celeste Sánchez SugíaBelum ada peringkat
- Dopaminergic Modulation of Short-Term Synaptic Plasticity at Striatal Inhibitory SynapsesDokumen6 halamanDopaminergic Modulation of Short-Term Synaptic Plasticity at Striatal Inhibitory SynapsesMaría Celeste Sánchez SugíaBelum ada peringkat
- Science:, 1870 (1999) Robert C. Malenka and and Roger A. NicollDokumen6 halamanScience:, 1870 (1999) Robert C. Malenka and and Roger A. NicollMaría Celeste Sánchez SugíaBelum ada peringkat
- Watson 2004Dokumen17 halamanWatson 2004María Celeste Sánchez SugíaBelum ada peringkat
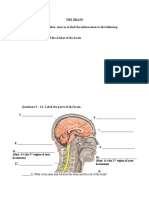
- Nervous System WorksheetDokumen3 halamanNervous System WorksheetaBelum ada peringkat
- Basal Ganglia-KreitzerDokumen51 halamanBasal Ganglia-KreitzerresearchphysioBelum ada peringkat
- Excitable Cells Lecture 2Dokumen34 halamanExcitable Cells Lecture 2stehephBelum ada peringkat
- NCM 106 - Pharmacology Midterms 1M-4M Edited PDFDokumen43 halamanNCM 106 - Pharmacology Midterms 1M-4M Edited PDFKisha Bethel100% (2)
- Brain Bee NotesDokumen10 halamanBrain Bee Notesapi-534972164Belum ada peringkat
- Reviewer On Nervous System Grade VIDokumen4 halamanReviewer On Nervous System Grade VIKent Francis LayaguinBelum ada peringkat
- The Brain: Learning & MemoryDokumen32 halamanThe Brain: Learning & MemoryAkanksha DubeyBelum ada peringkat
- Emotional Resilience - Part 1 The Evolving Brain PDFDokumen10 halamanEmotional Resilience - Part 1 The Evolving Brain PDFPeter Creagh67% (3)
- Neuroanesthesia and Cerebrospinal ProtectionDokumen697 halamanNeuroanesthesia and Cerebrospinal ProtectionUmi KalsumBelum ada peringkat
- Konsep Dasar Ilmu Biokimia Dan Biologi Molekuler Untuk Sistem SarafDokumen31 halamanKonsep Dasar Ilmu Biokimia Dan Biologi Molekuler Untuk Sistem SarafdiandraBelum ada peringkat
- MR IntracranIal menIngIomas!!! Vazno!!!Dokumen5 halamanMR IntracranIal menIngIomas!!! Vazno!!!StefanBelum ada peringkat
- Nervous System of FishDokumen4 halamanNervous System of Fishtalam_06100% (10)
- Roger Sperry: Pioneer of Neuronal Specificity: Essays On Aps Classic PapersDokumen3 halamanRoger Sperry: Pioneer of Neuronal Specificity: Essays On Aps Classic PapersAnderson HenriqueBelum ada peringkat
- Development of RhombencephalonDokumen28 halamanDevelopment of RhombencephalonRamesh KumarBelum ada peringkat
- Speech OutlineDokumen4 halamanSpeech Outlineapi-335645447Belum ada peringkat
- PersonalDevelopment - Q1 - Mod 5 - Powers-of-the-MindDokumen34 halamanPersonalDevelopment - Q1 - Mod 5 - Powers-of-the-MindGene RamirezBelum ada peringkat
- Study Guide Lin 2601Dokumen192 halamanStudy Guide Lin 2601Joanne100% (1)
- Control and Coordination - Class XDokumen5 halamanControl and Coordination - Class XMonika Mehan89% (19)
- SCIENCE ExamDokumen4 halamanSCIENCE ExamLesley AntojadoBelum ada peringkat
- Thalamic Stroke and BehaviourDokumen9 halamanThalamic Stroke and BehaviourparkdhingraBelum ada peringkat
- Grade 10 3rd Quarter Science ReviewerDokumen5 halamanGrade 10 3rd Quarter Science ReviewerZayin DelMundoBelum ada peringkat
- The Brain, Nervous System and Endocrine SystemDokumen13 halamanThe Brain, Nervous System and Endocrine SystemMark Lorenzo A RuizBelum ada peringkat
- Chapter 12 Coordination and ControlDokumen22 halamanChapter 12 Coordination and ControlXaundrae McDonaldBelum ada peringkat
- Disc Vis Lobe Front Et Tempo PDFDokumen12 halamanDisc Vis Lobe Front Et Tempo PDFMarina MaatoukBelum ada peringkat
- Connective TissueDokumen202 halamanConnective TissuePeter ThompsonBelum ada peringkat
- Development of The Nervous SystemDokumen27 halamanDevelopment of The Nervous Systemsalmankhan09215Belum ada peringkat
- Case Study On Meninomyocele (Repaired) PDokumen29 halamanCase Study On Meninomyocele (Repaired) Pcharanjit kaur100% (1)
- The Nervous SystemDokumen25 halamanThe Nervous SystemRoan Angela100% (4)
- SCI Pathophysiology MiculobDokumen1 halamanSCI Pathophysiology MiculobJemina GuenevereBelum ada peringkat
- Sci10 Q3 Module3Dokumen24 halamanSci10 Q3 Module3Jennifer Lomboy50% (2)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDDari EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDPenilaian: 5 dari 5 bintang5/5 (3)
- The Age of Magical Overthinking: Notes on Modern IrrationalityDari EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityPenilaian: 4 dari 5 bintang4/5 (30)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeDari EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BePenilaian: 2 dari 5 bintang2/5 (1)
- Love Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)Dari EverandLove Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)Penilaian: 3 dari 5 bintang3/5 (1)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionDari EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionPenilaian: 4 dari 5 bintang4/5 (404)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsDari EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsPenilaian: 4 dari 5 bintang4/5 (4)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisDari EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisPenilaian: 4.5 dari 5 bintang4.5/5 (42)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsDari EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsPenilaian: 5 dari 5 bintang5/5 (1)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsDari EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsBelum ada peringkat
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedDari EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedPenilaian: 5 dari 5 bintang5/5 (81)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsDari EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsPenilaian: 4.5 dari 5 bintang4.5/5 (170)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaDari EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Comfort of Crows: A Backyard YearDari EverandThe Comfort of Crows: A Backyard YearPenilaian: 4.5 dari 5 bintang4.5/5 (23)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Dari EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Penilaian: 4.5 dari 5 bintang4.5/5 (110)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDari EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityPenilaian: 4 dari 5 bintang4/5 (5)
- Summary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisDari EverandSummary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisPenilaian: 5 dari 5 bintang5/5 (8)
- Empath: The Survival Guide For Highly Sensitive People: Protect Yourself From Narcissists & Toxic Relationships. Discover How to Stop Absorbing Other People's PainDari EverandEmpath: The Survival Guide For Highly Sensitive People: Protect Yourself From Narcissists & Toxic Relationships. Discover How to Stop Absorbing Other People's PainPenilaian: 4 dari 5 bintang4/5 (95)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeDari EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifePenilaian: 4.5 dari 5 bintang4.5/5 (253)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessDari EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessPenilaian: 4.5 dari 5 bintang4.5/5 (328)
- Summary: Thinking, Fast and Slow: by Daniel Kahneman: Key Takeaways, Summary & Analysis IncludedDari EverandSummary: Thinking, Fast and Slow: by Daniel Kahneman: Key Takeaways, Summary & Analysis IncludedPenilaian: 4 dari 5 bintang4/5 (61)
- The Obesity Code: Unlocking the Secrets of Weight LossDari EverandThe Obesity Code: Unlocking the Secrets of Weight LossPenilaian: 4 dari 5 bintang4/5 (6)
- The Marshmallow Test: Mastering Self-ControlDari EverandThe Marshmallow Test: Mastering Self-ControlPenilaian: 4.5 dari 5 bintang4.5/5 (60)
- Manipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesDari EverandManipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesPenilaian: 4.5 dari 5 bintang4.5/5 (1412)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryDari EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryPenilaian: 4 dari 5 bintang4/5 (45)