Anda mungkin juga menyukai
- Polycell Evacell: For A Multitude of UsesDokumen7 halamanPolycell Evacell: For A Multitude of UsesJean Carlos Arias SaldañaBelum ada peringkat
- Polycell Evacell: For A Multitude of UsesDokumen7 halamanPolycell Evacell: For A Multitude of UsesJean Carlos Arias SaldañaBelum ada peringkat
- ALP - Basic 2Dokumen2 halamanALP - Basic 2Jean Carlos Arias SaldañaBelum ada peringkat
- ALP - Basic 2Dokumen2 halamanALP - Basic 2Jean Carlos Arias SaldañaBelum ada peringkat
- Taylor - Encouraging Industry To Assess and Implement Cleaner ProductionDokumen9 halamanTaylor - Encouraging Industry To Assess and Implement Cleaner ProductionJean Carlos Arias SaldañaBelum ada peringkat
- Khan - Cleaner ProductionDokumen6 halamanKhan - Cleaner ProductionJean Carlos Arias SaldañaBelum ada peringkat
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Biology Form 4 Compilation of ExperimentsDokumen28 halamanBiology Form 4 Compilation of Experimentsriyashree50% (4)
- 2 5364059534434239298Dokumen182 halaman2 5364059534434239298RasoulBelum ada peringkat
- Chapter 2 LipidsDokumen109 halamanChapter 2 LipidsTiffany Shane VallenteBelum ada peringkat
- Waste Management - A PerspectiveDokumen6 halamanWaste Management - A PerspectiveInternational Journal of Innovative Science and Research TechnologyBelum ada peringkat
- Formulated Solvent Reduces Shale Gas Processing Costs - PTQ - Gas 2014Dokumen6 halamanFormulated Solvent Reduces Shale Gas Processing Costs - PTQ - Gas 2014onizuka-t2263Belum ada peringkat
- Quality Assurance and Quality Control in Environmental AnalysisDokumen97 halamanQuality Assurance and Quality Control in Environmental Analysisantolin beceril100% (1)
- A01 425Dokumen23 halamanA01 425jaimeBelum ada peringkat
- EIA For Parind HPPDokumen17 halamanEIA For Parind HPPLawi HPP, ACE PeshawarBelum ada peringkat
- DNA Extraction OverviewDokumen14 halamanDNA Extraction OverviewNicholas SoBelum ada peringkat
- CHEMISTRY F3 SET 01 May 2020 PDFDokumen3 halamanCHEMISTRY F3 SET 01 May 2020 PDFumuchero danielBelum ada peringkat
- 1 s2.0 B9780444634283501260 MainDokumen6 halaman1 s2.0 B9780444634283501260 MainDavid ManningBelum ada peringkat
- Bang Tra Hoa Ly 1Dokumen42 halamanBang Tra Hoa Ly 1Hoài ThươngBelum ada peringkat
- RA 6969 (Permitting)Dokumen84 halamanRA 6969 (Permitting)Christine Dhianne FloresBelum ada peringkat
- ST 5730 2016 Add 9 enDokumen199 halamanST 5730 2016 Add 9 enbi2458Belum ada peringkat
- Pengendalian Mikroba DG Hurdle TechDokumen12 halamanPengendalian Mikroba DG Hurdle TechetikaBelum ada peringkat
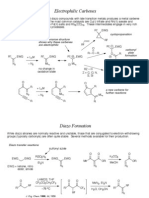
- Carbenoids 2Dokumen15 halamanCarbenoids 2costea0028Belum ada peringkat
- The DoorDokumen3 halamanThe DoorAliah Necole EsquibelBelum ada peringkat
- EvaluationofCalotropisgigantea - ComposMater09Dokumen10 halamanEvaluationofCalotropisgigantea - ComposMater09FrankensteinBelum ada peringkat
- Lab 2 - Identification of Functional GroupsDokumen14 halamanLab 2 - Identification of Functional GroupsShamaya Murray60% (5)
- PRODUCTIONOF60000MTPAOFOLEOCHEMICALMETHYLESTERFROMRBDPALMKERNELOILDokumen586 halamanPRODUCTIONOF60000MTPAOFOLEOCHEMICALMETHYLESTERFROMRBDPALMKERNELOILKevin Fernando PratamaBelum ada peringkat
- Hyperconjugation - Chemistry LibreTextsDokumen2 halamanHyperconjugation - Chemistry LibreTextsAatmaanandaBelum ada peringkat
- SDS - Clarity Synthetic EA Grease SeriesDokumen9 halamanSDS - Clarity Synthetic EA Grease SeriesGeorgina SuleBelum ada peringkat
- Carbonyl ChemsketchDokumen3 halamanCarbonyl ChemsketchJenica RiaBelum ada peringkat
- Diallo 2016Dokumen11 halamanDiallo 2016Dương Minh MẫnBelum ada peringkat
- (Food Safety) Table OlivesDokumen32 halaman(Food Safety) Table OlivesChiêu Nghi VõBelum ada peringkat
- Synthesis of Near-Infrared Heptamethine Cyanine DyesDokumen132 halamanSynthesis of Near-Infrared Heptamethine Cyanine DyesNgọc Quyền PhạmBelum ada peringkat
- Acrylonitrile-Butadiene-Styrene (ABS) Market Analysis & Forecast To 2022Dokumen2 halamanAcrylonitrile-Butadiene-Styrene (ABS) Market Analysis & Forecast To 2022Mahesh ChaudhariBelum ada peringkat
- Alkaloids-: Nitrogen Terresterial Plants Spectrum Physiological Studies Synthetic DrugsDokumen21 halamanAlkaloids-: Nitrogen Terresterial Plants Spectrum Physiological Studies Synthetic DrugsAnuj KaundalBelum ada peringkat
- Organic Chemistry Cambridge As-Level SummaryDokumen4 halamanOrganic Chemistry Cambridge As-Level SummaryReynaldi BozzBelum ada peringkat
- Electrical Conductivity of Aqueous SolutionsDokumen1 halamanElectrical Conductivity of Aqueous SolutionslaughingalirezaBelum ada peringkat